
Integration of Whole-Genome Resequencing and Transcriptome Sequencing Reveals Candidate Genes in High Glossiness of Eggshell
 and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
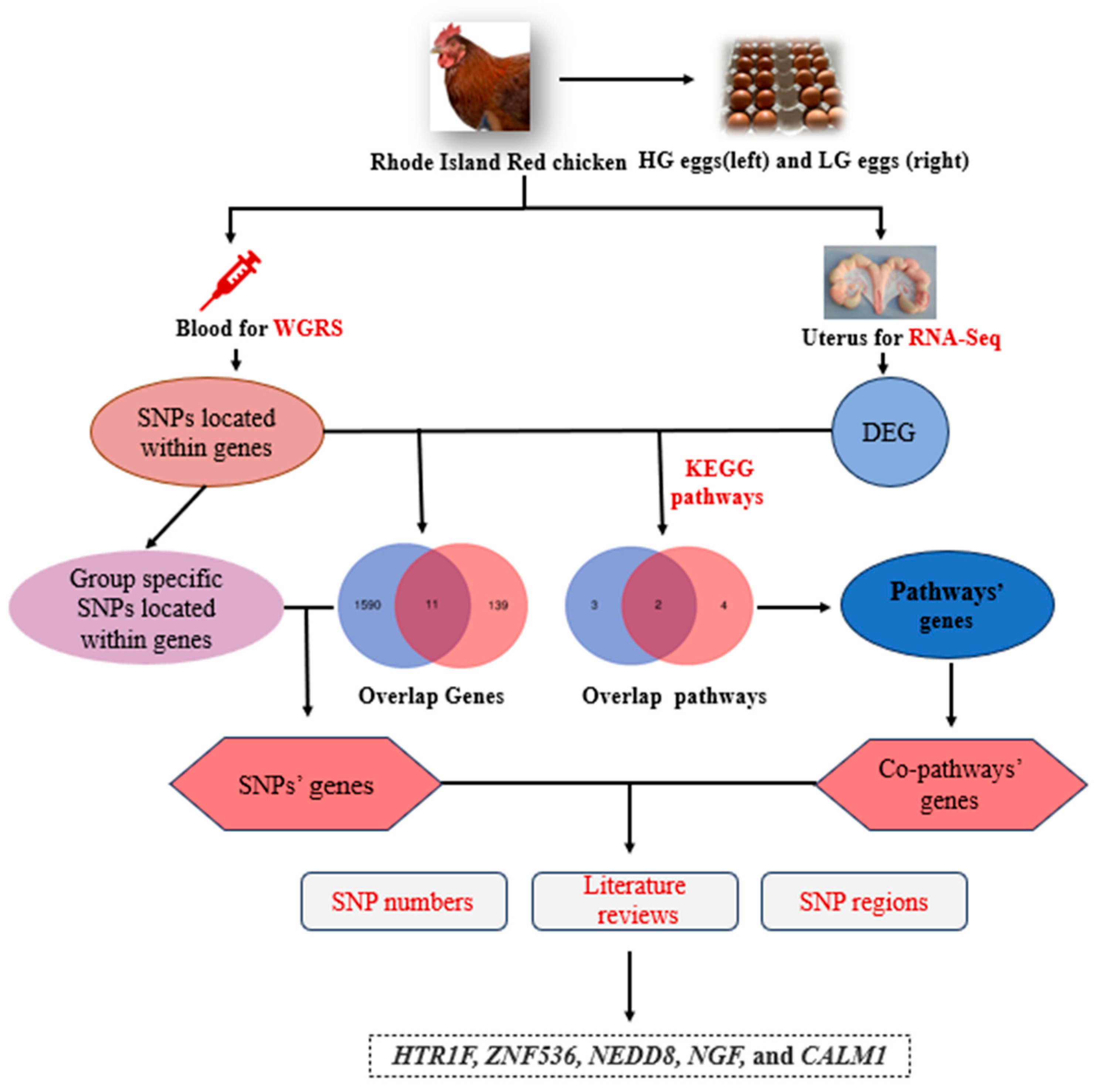
2.1. Birds and Sample Collection
2.2. Eggshell Quality Measurement
2.3. Genomic DNA Extraction, RNA Extraction and qPCR Assay
2.4. Transcriptome Sequencing (RNA-seq)
2.5. Whole-Genome Resequencing
2.6. Enrichment Analysis
2.7. Data Analysis
3. Results
3.1. Measurement and Analysis of Eggshell Gloss
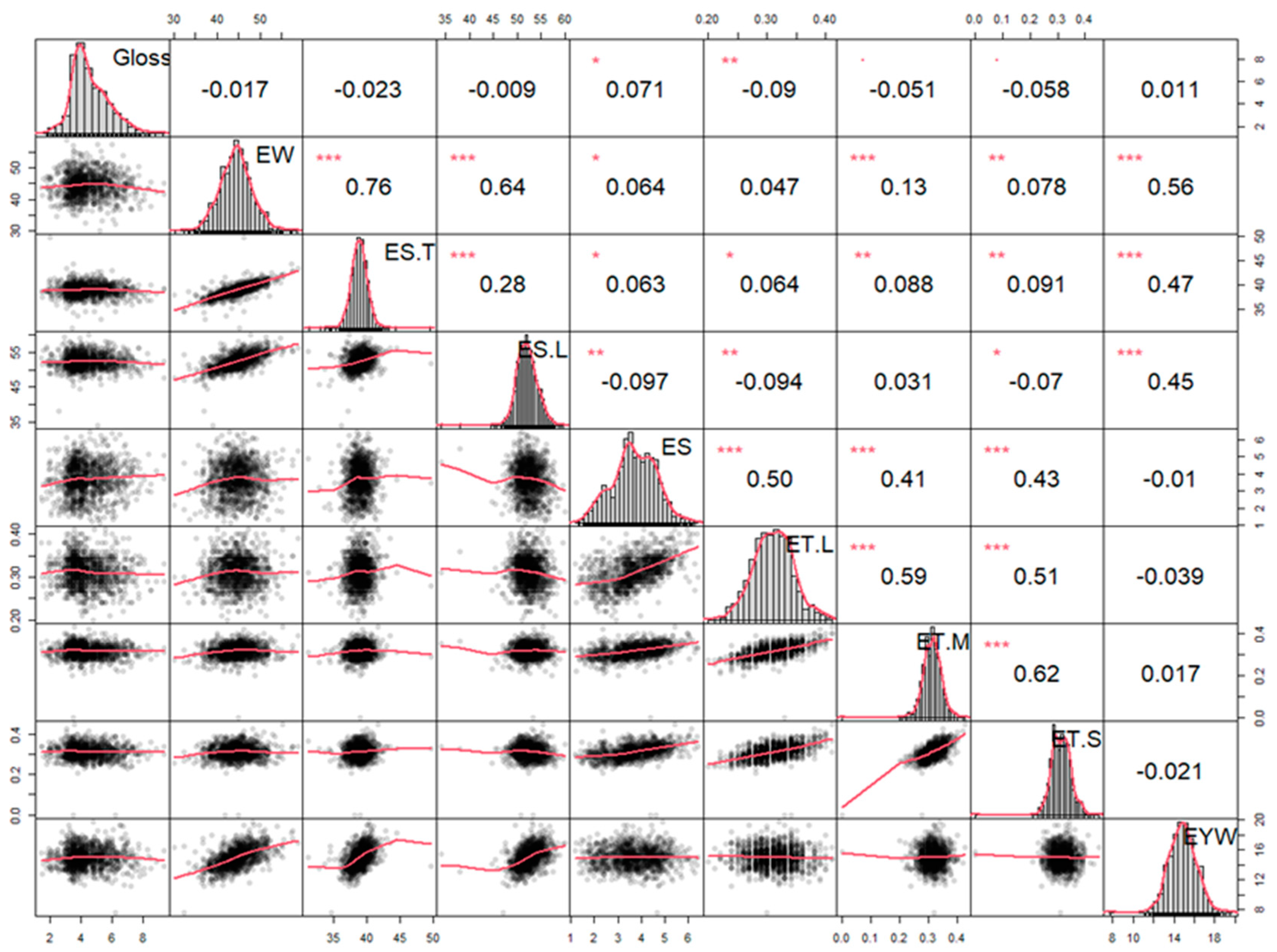
3.2. Population Glossiness Distribution and Correlation Analysis
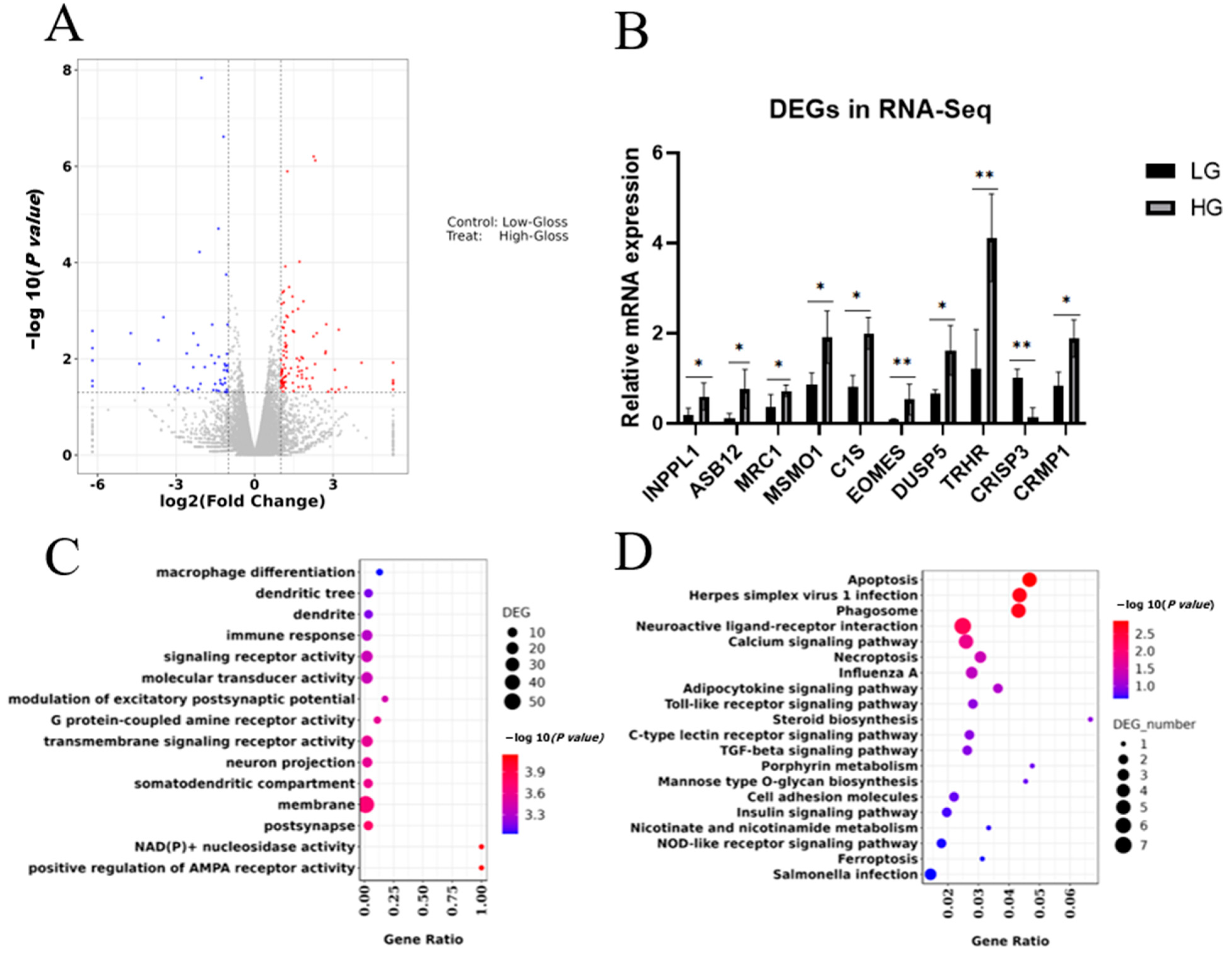
3.3. RNA-seq Revealed Significant Differences in Gene Expression Patterns Related to Eggshell Gloss
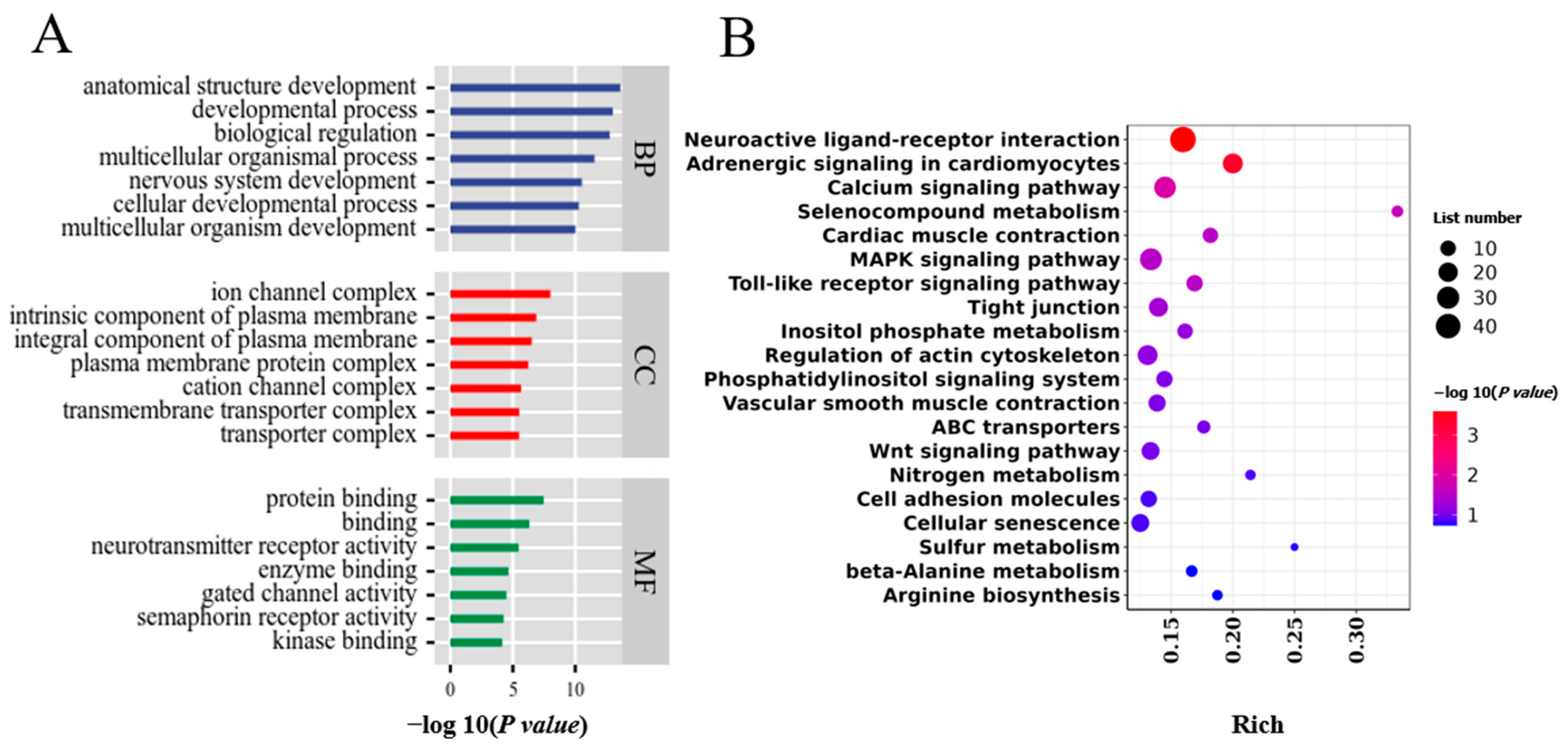
3.4. WGRS Analysis to Identify Group-Specific Genetic Variation and Associated Genes
3.5. Joint Analysis of WGRS and RNA-seq
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| WGRS | whole-genome resequencing |
| RNA-seq | transcriptome sequencing |
| HG | high gloss |
| LG | low gloss |
| DEG | differentially expressed gene |
| SEM | scanning electron microscope |
| GO | Gene Ontology |
| mRNA | messenger RNA |
| qPCR | quantitative real-time PCR |
| BP | biological process |
| CC | cellular component |
| MF | molecular function |
| bp | base pair |
| WGCNA | weighted correlation network analysis |
References
- Gautron, J.; Dombre, C.; Nau, F.; Feidt, C.; Guillier, L. Review: Production factors affecting the quality of chicken table eggs and egg products in Europe. Animal 2022, 16, 100425. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ren, J.; Wang, K.; Zheng, J.; Xu, G.; Ge, C.; Yang, N.; Sun, C. The effect of breed and age on the gloss of chicken eggshells. Poult. Sci. 2020, 99, 2494–2499. [Google Scholar] [CrossRef] [PubMed]
- Ketta, M.; Tůmová, E. Eggshell structure, measurements, and quality-affecting factors in laying hens: A review. Czech J. Anim. Sci. 2016, 61, 299–309. [Google Scholar] [CrossRef]
- Chadwick, A.C.; Kentridge, R.W. The perception of gloss: A review. Vis. Res. 2015, 109, 221–235. [Google Scholar] [CrossRef]
- Trezza, T.A.; Krochta, J.M. Specular reflection, gloss, roughness and surface heterogeneity of biopolymer coatings. J. Appl. Polym. Sci. 2001, 79, 2221–2229. [Google Scholar] [CrossRef]
- Igic, B.; Fecheyr-Lippens, D.; Xiao, M.; Chan, A.; Hanley, D.; Brennan, P.R.; Grim, T.; Waterhouse, G.I.; Hauber, M.E.; Shawkey, M.D. A nanostructural basis for gloss of avian eggshells. J. R. Soc. Interface 2015, 12, 20141210. [Google Scholar] [CrossRef]
- Zhang, X.; Li, G.; Duan, Z.; Li, H.; Yan, Y.; Han, W.; Wu, G. Quantitative measurement and statistical analysis of egg shell gloss. Heilongjiang Anim. Sci. Vet. Med. 2021, 7, 67–70. [Google Scholar] [CrossRef]
- Wilson, N.; Preston, E.J.; Preston, F.W. The Gloss of Eggs. Auk 1958, 75, 456–464. [Google Scholar] [CrossRef]
- Li, Q.; Wang, K.; Zheng, J.; Sun, C.; Ge, C.; Yang, N.; Xu, G. Nanostructural basis for the gloss of chicken eggshells. Poult. Sci. 2019, 98, 5446–5451. [Google Scholar] [CrossRef]
- Grizzle, J.; Iheanacho, M.; Saxton, A.; Broaden, J. Nutritional and environmental factors involved in egg shell quality of laying hens. Br. Poult. Sci. 1992, 33, 781–794. [Google Scholar] [CrossRef]
- Liu, L.; Fan, Y.; Zhang, Z.; Yang, C.; Geng, T.; Gong, D.; Hou, Z.; Ning, Z. Analysis of gene expression and regulation implicates C2H9orf152 has an important role in calcium metabolism and chicken reproduction. Anim. Reprod. Sci. 2017, 176, 1–10. [Google Scholar] [CrossRef]
- Fan, Y.F.; Hou, Z.C.; Yi, G.Q.; Xu, G.Y.; Yang, N. The sodium channel gene family is specifically expressed in hen uterus and associated with eggshell quality traits. BMC Genet. 2013, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Brionne, A.; Nys, Y.; Hennequet-Antier, C.; Gautron, J. Hen uterine gene expression profiling during eggshell formation reveals putative proteins involved in the supply of minerals or in the shell mineralization process. BMC Genom. 2014, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Jonchère, V.; Réhault-Godbert, S.; Hennequet-Antier, C.; Cabau, C.; Sibut, V.; Cogburn, L.A.; Nys, Y.; Gautron, J. Gene expression profiling to identify eggshell proteins involved in physical defense of the chicken egg. BMC Genom. 2010, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Ying, F.; Li, Q.; Zhang, G.; Zhang, J.; Liu, R.; Zheng, M.; Wen, J.; Zhao, G. A significant quantitative trait locus on chromosome Z and its impact on egg production traits in seven maternal lines of meat-type chicken. J. Anim. Sci. Biotechnol. 2022, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Shi, Z.; Ou, X.; Liu, G. Whole-genome resequencing reveals genetic indels of feathered-leg traits in domestic chickens. J. Genet. 2019, 98, 47. [Google Scholar] [CrossRef] [PubMed]
- Geibel, J.; Praefke, N.P.; Weigend, S.; Simianer, H.; Reimer, C. Assessment of linkage disequilibrium patterns between structural variants and single nucleotide polymorphisms in three commercial chicken populations. BMC Genom. 2022, 23, 193. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Putri, G.H.; Anders, S.; Pyl, P.T.; Pimanda, J.E.; Zanini, F. Analysing high-throughput sequencing data in Python with HTSeq 2.0. Bioinformatics 2022, 38, 2943–2945. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Volume 1. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; He, L.; Li, Y.; Huang, W.; Xi, F.; Lin, L.; Zhi, Q.; Zhang, W.; Tang, Y.T.; Geng, C.; et al. OTG-snpcaller: An optimized pipeline based on TMAP and GATK for SNP calling from ion torrent data. PLoS ONE 2014, 9, e97507. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; Guo, J.R.; Yang, H.M.; Zhou, R.J.; Liu, J.X.; Li, S.Z.; Dong, C.Y. Differential expression profiling of ovarian genes in prelaying and laying geese. Poult. Sci. 2009, 88, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Jesús, M.; Avilés, J.; Stokke, B.; Moksnes, A.; Røskaft, E.; Moller, A. Nest predation and the evolution of egg appearance in passerine birds in Europe and North America. Evol. Ecol. Res. 2006, 8, 493–513. [Google Scholar]
- Hanley, D.; Stoddard, M.C.; Cassey, P.; Brennan, P.L.R. Eggshell Conspicuousness in Ground Nesting Birds: Do Conspicuous Eggshells Signal Nest Location to Conspecifics? Avian Biol. Res. 2013, 6, 147–156. [Google Scholar] [CrossRef]
- Bello, S.F.; Xu, H.; Guo, L.; Li, K.; Zheng, M.; Xu, Y.; Zhang, S.; Bekele, E.J.; Bahareldin, A.A.; Zhu, W.; et al. Hypothalamic and ovarian transcriptome profiling reveals potential candidate genes in low and high egg production of white Muscovy ducks (Cairina moschata). Poult. Sci. 2021, 100, 101310. [Google Scholar] [CrossRef]
- Nys, Y.; Le Roy, N. Chapter 22—Calcium Homeostasis and Eggshell Biomineralization in Female Chicken. In Vitamin D, 4th ed.; Feldman, D., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 361–382. [Google Scholar]
- Zhang, Y.; Wang, S.; Ruan, D.; Zheng, C. Calcium Metabolism Mechanism of Eggshell Formation and lts Influencing Factors. Chin. J. Anim. Nutr. 2021, 33, 1201–1207. [Google Scholar]
- Huang, C.; Yue, Q.; Hao, E.; Xu, Y.; Zhao, X.; Dai, Z.; Chen, H. Effects of Dietary Calcium Level on Production Performance, Eqg Quality and Serum Biochemica Parameters of Hens during the Postpeak Period. China Poult. 2018, 40, 25–29. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, K.; Feng, B.; Zhang, Z.; Wang, R.; Tang, L.; Li, W.; Li, Q.; Piferrer, F.; Shao, C. Transcriptome of Gonads From High Temperature Induced Sex Reversal During Sex Determination and Differentiation in Chinese Tongue Sole, Cynoglossus semilaevis. Front. Genet. 2019, 10, 1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, L.; Han, K.; Zhang, X.; Zhang, G.; Dai, G.; Wang, J.; Xie, K. Transcriptome analysis of ovary in relatively greater and lesser egg producing Jinghai Yellow Chicken. Anim. Reprod. Sci. 2019, 208, 106114. [Google Scholar] [CrossRef] [PubMed]
- Mu, R.; Yu, Y.-y.; Gegen, T.; Wen, D.; Wang, F.; Chen, Z.; Xu, W.-b. Transcriptome analysis of ovary tissues from low- and high-yielding Changshun green-shell laying hens. BMC Genom. 2021, 22, 349. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liu, J.; Cao, J.; Zhang, H.; Liu, X.J.G. Ovarian transcriptomic analysis of black Muscovy duck at the early, peak and late egg-laying stages. Gene 2021, 777, 145449. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, W.; Zhang, H.X.; Ruan, D.; Lin, Y.C. Influence of particle size and calcium source on production performance, egg quality, and bone parameters in laying ducks. Poult. Sci. 2014, 93, 2560–2566. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.W.; Qi, G.H.; Cui, C.F.; Wu, S.G.; Zhang, H.J.; Xu, L.; Wang, J. Effects of dietary Bacillus subtilis supplementation and calcium levels on performance and eggshell quality of laying hens in the late phase of production. Poult. Sci. 2021, 100, 100970. [Google Scholar] [CrossRef] [PubMed]
- Dijkslag, M.A.; Kwakkel, R.P.; Martin-Chaves, E.; Alfonso-Carrillo, C.; Walvoort, C.; Navarro-Villa, A. The effects of dietary calcium and phosphorus level, and feed form during rearing on growth performance, bone traits and egg production in brown egg-type pullets from 0 to 32 weeks of age. Poult. Sci. 2021, 100, 101130. [Google Scholar] [CrossRef]
- Kupittayanant, S.; Kupittayanant, P.; Suwannachat, C. Mechanisms of uterine contractility in laying hens. Anim. Reprod. Sci. 2009, 115, 215–224. [Google Scholar] [CrossRef]
- Levorse, J.M.; Tilly, J.L.; Johnson, A.L. Role of calcium in the regulation of theca cell androstenedione production in the domestic hen. J. Reprod. Fertil. 1991, 92, 159–167. [Google Scholar] [CrossRef][Green Version]
- Chin, D.; Means, A.R. Calmodulin: A prototypical calcium sensor. Trends Cell Biol. 2000, 10, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Bai, Y.; Zhang, Q.; Zhang, H.; Fan, Y.; Han, H.; Liu, Y. Associations of CALM1 and DRD1 polymorphisms, and their expression levels, with Taihang chicken egg-production traits. Anim. Biotechnol. 2023, 34, 994–1004. [Google Scholar] [CrossRef]
- Xu, Q.; He, H.; Ma, R.; Chen, Z.; Yi, Z.; Kang, B.; Jiang, D. Characterization and Differential Expression Profile of Calm1 Gene in Sichuan White Goose. Acta Agric. Boreali-Sin. 2016, 31, 128–133. [Google Scholar]
- Li, J.; Wu, L.; Tan, G.; Qin, Y.; Zhang, Y. ldentification and Analysis of SNPs of Eggshell Quality Related CALM1 Gene Mutation in Sansui Duck. J. Mt. Agric. Biol. 2020, 39, 67–72. [Google Scholar] [CrossRef]
- Sun, C.; Qiu, Y.; Ren, Q.; Zhang, X.; Cao, B.; Zou, Y.; Li, J.; Zhang, J.; Wang, Y. Molecular Cloning and Functional Characterization of Three 5-HT Receptor Genes (HTR1B, HTR1E, and HTR1F) in Chickens. Genes 2021, 12, 891. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.; Hu, S.; Wang, G.; Hu, J.; Zhang, J.; Li, L.; Hu, B.; He, H.; Liu, H.; Xia, L.; et al. Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production. Genes 2020, 11, 455. [Google Scholar] [CrossRef]
- Liu, W.; Li, D.; Liu, J.; Chen, S.; Qu, L.; Zheng, J.; Xu, G.; Yang, N. A genome-wide SNP scan reveals novel loci for egg production and quality traits in white leghorn and brown-egg dwarf layers. PLoS ONE 2011, 6, e28600. [Google Scholar] [CrossRef]
- Mori, F.; Nishie, M.; Piao, Y.-S.; Kito, K.; Kamitani, T.; Takahashi, H.; Wakabayashi, K. Accumulation of NEDD8 in neuronal and glial inclusions of neurodegenerative disorders. Neuropathol. Appl. Neurobiol. 2005, 31, 53–61. [Google Scholar] [CrossRef]
- Ma, X.; Guo, X.; Jia, K.; Cheng, Y.; Lu, L. Preliminary Study on Mechanism of bta-miR-1343-3p Targeting NEDD8Regulating CellProliferation and Apoptosis of Bovine Granulosa Cells. J. Shanxi Agric. Sci. 2023, 51, 198–205. [Google Scholar]
- Denk, F.; Bennett, D.L.; McMahon, S.B. Nerve Growth Factor and Pain Mechanisms. Annu. Rev. Neurosci. 2017, 40, 307–325. [Google Scholar] [CrossRef]
- Antipova, T.; Gudasheva, T.; Tarasiuk, A.; Nikolajev, S.; Logvinov, I.; Kruglov, S.; Seredenin, S. Effects of new low-molecular mimetic of NGF loop 3 on the activation of TrkA-receptor, PI3K/AKT and MAPK/ERK signaling pathways. Eur. Neuropsychopharmacol. 2016, 26, S228–S229. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | log2FoldChange | p Value | Chr | Regulation (HG vs. LG) |
|---|---|---|---|---|
| FGF19 | −2.03066 | 1.45 × 10−8 | 5 | Down |
| SOUL | −1.18806 | 2.43 × 10−7 | 5 | Down |
| ENSGALG00000031427 | 2.258618 | 6.25 × 10−7 | 3 | Up |
| ENSGALG00000009479 | 2.318946 | 7.55 × 10−7 | 2 | Up |
| C1S | 1.2519 | 1.27 × 10−6 | 1 | Up |
| APOD | −1.3807 | 1.97 × 10−5 | 9 | Down |
| ZNF536 | −2.10433 | 6.03 × 10−5 | 11 | Down |
| TRHR | 1.716454 | 9.61 × 10−5 | 2 | Up |
| AQP1 | 1.173698 | 0.00012 | 2 | Up |
| PTGFRN | −1.0838 | 0.000178 | 1 | Down |
| Region | Number | Percentage |
|---|---|---|
| Intronic | 8125 | 55.37% |
| Intergenic | 5813 | 39.62% |
| Exonic | 186 | 1.27% |
| Splicing | 2 | 0.01% |
| Downstream | 110 | 0.75% |
| Upstream | 98 | 0.67% |
| Upstream/downstream | 13 | 0.09% |
| UTR5 | 63 | 0.43% |
| UTR3 | 261 | 1.78% |
| UTR5/UTR3 | 2 | 0.01% |
| Total | 15,040 | 100% |
| Gene Name | p Value | log2FoldChange | Chr | SNP Num |
|---|---|---|---|---|
| ZNF536 | 6.03 × 10−5 | −2.104328726 | 11 | 20 |
| CRMP1 | 0.000392 | 1.115834887 | 4 | 1 |
| LYVE1 | 0.005761 | 1.194353931 | 5 | 3 |
| CNTN1 | 0.016508 | 1.049463725 | 1 | 7 |
| NOX3 | 0.014941 | −2.343634297 | 3 | 3 |
| ARHGAP15 | 0.020556 | 1.04431234 | 7 | 1 |
| INPPL1 | 0.022631 | 1.6402217 | 4 | 1 |
| HTR1F | 0.031011 | Inf | 1 | 2 |
| MRC1 | 0.040211 | 1.005384815 | 2 | 1 |
| C17orf58 | 0.043432 | −1.064341843 | 18 | 4 |
| TFEC | 0.048114 | 2.81415403 | 1 | 1 |
| Co-Pathways | Calcium Signaling Pathway | Neuroactive Ligand–Receptor Interaction |
|---|---|---|
| RNA-seq | TACR3, TRHR, CD38, CHRNA8, FGF19 | HTR1F, TACR3, GRM4, CHRNA8 |
| WGRS | NGF, CALM1, PLCD1, NOS2, STIM2, ADCY9, CACNA1D, GRIN2A, PPP3R1, CHRM2, FGF3, CACNA1B, ERBB3, HTR7, CHRM3, CAMK4, ADRA1B, NTSR1, PLCB1, HTR2C, FGF18, ERBB4, ATP2B1, SLC8A3, FGF10, CHRM5, GNAL, CCKAR | HTR1F, RLN3, GRID1, VIPR2, DRD2, PARD3, GLRA2, ADRA2A, HCRTR2, CHRNA5, FSHR, GRIK4, PTH2R, HTR7, CHRM3, NTSR1, THRB, GABRA4, SSTR4, HTR2C, CCKAR, GRIN2A, CHRM2, MC4R, GABRQ, SSTR5, ADCYAP1R1, GRM3, ADRA1B, GABRB3, CALCRL, GRID2, LPAR4, GLP1R, CHRM5, GLRB |
| Gene | Site | Chr | Region | Base in HG | Base in LG |
|---|---|---|---|---|---|
| NEDD8 | 506704 | 35 | exonic | C | T |
| 506810 | 35 | exonic | C | T | |
| 505577 | 35 | exonic | G | A | |
| NGF | 4069359 | 26 | UTR5 | A | G |
| 4070415 | 26 | UTR5 | C | T | |
| HTR1F | 92728689 | 1 | intergenic | T | C |
| 92970994 | 1 | intergenic | T | C | |
| ZNF536 | 8393350 | 11 | intronic | C | G |
| 8583435 | 11 | intronic | A | G | |
| 8591108 | 11 | intronic | G | A | |
| CALM1 | 43490289 | 5 | intergenic | G | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Li, S.; He, S.; Zheng, H.; Li, R.; Liu, L.; Geng, T.; Zhao, M.; Gong, D. Integration of Whole-Genome Resequencing and Transcriptome Sequencing Reveals Candidate Genes in High Glossiness of Eggshell. Animals 2024, 14, 1141. https://doi.org/10.3390/ani14081141
Song X, Li S, He S, Zheng H, Li R, Liu L, Geng T, Zhao M, Gong D. Integration of Whole-Genome Resequencing and Transcriptome Sequencing Reveals Candidate Genes in High Glossiness of Eggshell. Animals. 2024; 14(8):1141. https://doi.org/10.3390/ani14081141
Chicago/Turabian StyleSong, Xiang, Shuo Li, Shixiong He, Hongxiang Zheng, Ruijie Li, Long Liu, Tuoyu Geng, Minmeng Zhao, and Daoqing Gong. 2024. "Integration of Whole-Genome Resequencing and Transcriptome Sequencing Reveals Candidate Genes in High Glossiness of Eggshell" Animals 14, no. 8: 1141. https://doi.org/10.3390/ani14081141
APA StyleSong, X., Li, S., He, S., Zheng, H., Li, R., Liu, L., Geng, T., Zhao, M., & Gong, D. (2024). Integration of Whole-Genome Resequencing and Transcriptome Sequencing Reveals Candidate Genes in High Glossiness of Eggshell. Animals, 14(8), 1141. https://doi.org/10.3390/ani14081141