
A Potential “Vitaminic Strategy” against Caries and Halitosis

 , , , ,
, , , ,  and
and _Di_Marco.png)
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Effect of Vea® Oris Treatments on Planktonic and Biofilm Growth
2.2. Potential Ability of Vea® Oris Treatments to Modulate H2S Production
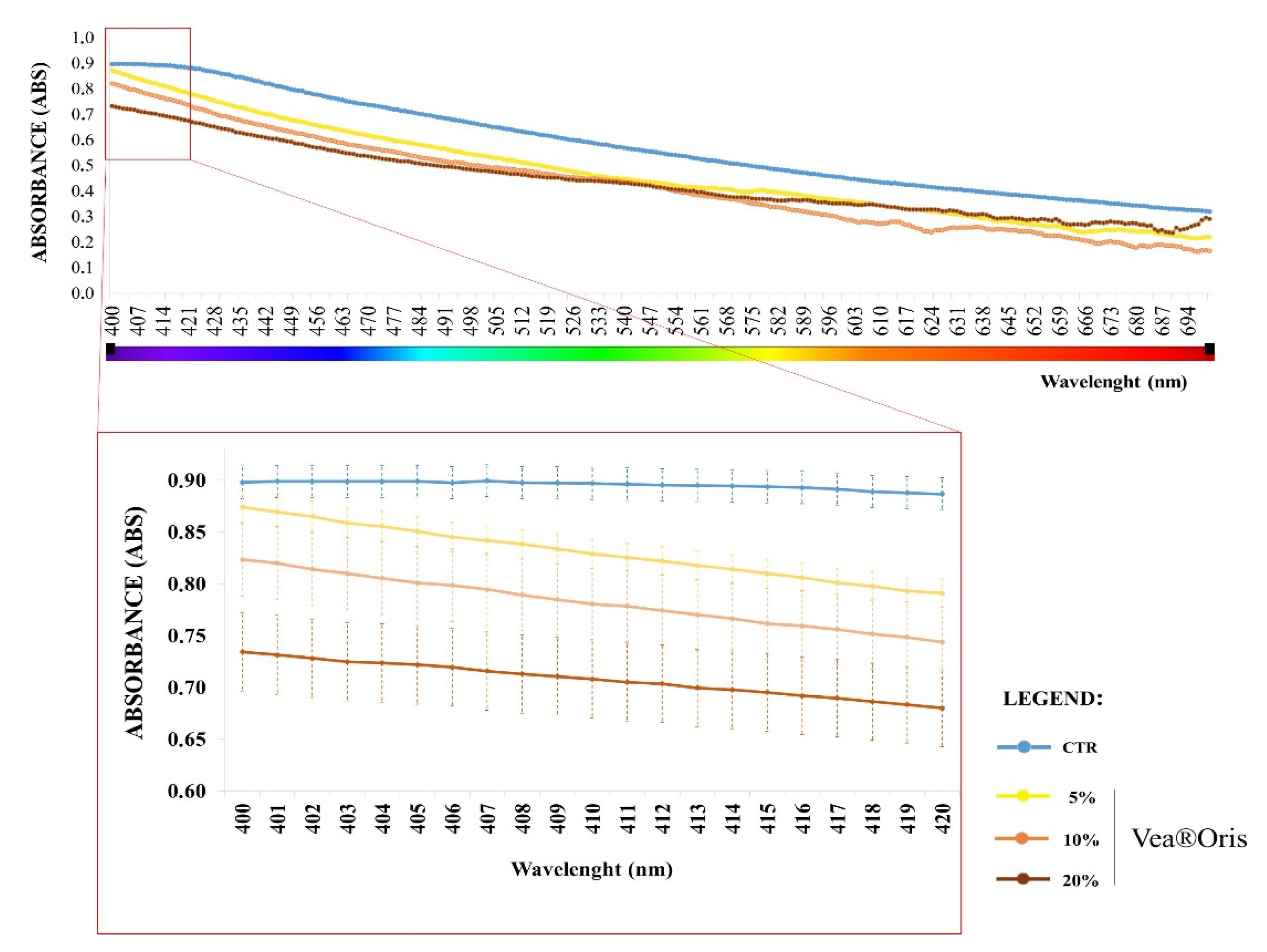
3. Results
3.1. Variation of Planktonic and Biofilm Growth
3.2. Variation of H2S Production
4. Discussion
5. Conclusions
Limitations of the Study
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, M.; Xuan, S.; Wang, Z. Oral microbiota: A new view of body health. Food Sci. Hum. Wellness 2019, 8, 8–15. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome-An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.J., Jr. Composition and development of oral bacterial communities. Periodontology 2000 2014, 64, 20–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo Ulloa, P.; van der Veen, M.H.; Krom, B.P. Review: Modulation of the oral microbiome by the host to promote ecological balance. Odontology 2019, 107, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Willis, J.R.; Gabaldón, T. The human oral microbiome in health and disease: From sequences to ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Shokeen, B.; He, X.; Shi, W.; Lux, R. Streptococcus mutans SpaP binds to RadD of Fusobacterium nucleatum ssp. polymorphum. Mol. Oral Microbiol. 2017, 32, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Gunsolley, J. Application of metagenomics in understanding oral health and disease. Virulence 2014, 5, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Belda-Ferre, P.; Alcaraz, L.D.; Cabrera-Rubio, R.; Romero, H.; Simón-Soro, A.; Pignatelli, M.; Mira, A. The oral metagenome in health and disease. ISME J. 2012, 6, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [Green Version]
- Harper, R.A.; Carpenter, G.H.; Proctor, G.B.; Harvey, R.D.; Gambogi, R.J.; Geonnotti, A.R.; Hider, R.; Jones, S.A. Diminishing biofilm resistance to antimicrobial nanomaterials through electrolyte screening of electrostatic interactions. Colloids Surf. B Biointerfaces 2019, 173, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Jwa, S.K. Efficacy of moringa oleifera leaf extracts against cariogenic biofilm. Prev. Nutr. Food Sci. 2019, 24, 308–312. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Belda-Ferre, P.; Simón-Soro, A.; Mira, A. Microbiota diversity and gene expression dynamics in human oral biofilms. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thurnheer, T.; Karygianni, L.; Flury, M.; Belibasakis, G.N. Fusobacterium Species and Subspecies Differentially Affect the Composition and Architecture of Supra- and Subgingival Biofilms Models. Front. Microbiol. 2019, 10, 1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karched, M.; Bhardwaj, R.G.; Asikainen, S.E. Coaggregation and biofilm growth of Granulicatella spp. with Fusobacterium nucleatum and Aggregatibacter actinomycetemcomitans. BMC Microbiol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenartova, M.; Tesinska, B.; Janatova, T.; Hrebicek, O.; Mysak, J.; Janata, J.; Najmanova, L. The Oral Microbiome in Periodontal Health. Front. Cell. Infect. Microbiol. 2021, 11, 629723. [Google Scholar] [CrossRef]
- Saputri, D.; Abrar, M.; Mubarak, Z.; Mudatsir. The Role of Fusobacterium Nucleatum on Chronic Periodontitis (Literature Review). Adv. Health Sci. Res. 2021, 32, 17–21. [Google Scholar] [CrossRef]
- Washio, J.; Sato, T.; Koseki, T.; Takahashi, N. Hydrogen sulfide-producing bacteria in tongue biofilm and their relationship with oral malodour. J. Med. Microbiol. 2005, 54, 889–895. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Nakano, Y.; Watanabe, T.; Yoneda, M.; Hirofuji, T.; Hanioka, T. Two mechanisms of oral malodor inhibition by zinc ions. J. Appl. Oral Sci. 2018, 26, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kuang, X.; Chen, V.; Xu, X. Novel Approaches to the Control of Oral Microbial Biofilms. Biomed. Res. Int. 2018, 2018, 6498932. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Yoshida, A.; Awano, S.; Ansai, T.; Takehara, T. Quantitative detection of volatile sulfur compound-producing microorganisms in oral specimens using real-time PCR. Oral Dis. 2005, 11, 67–71. [Google Scholar] [CrossRef]
- Yoshida, A.; Yoshimura, M.; Ohara, N.; Yoshimura, S.; Nagashima, S.; Takehara, T.; Nakayama, K. Hydrogen Sulfide Production From Cysteine and Homocysteine by Periodontal and Oral Bacteria. J. Periodontol. 2009, 80, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- Verspecht, T.; Rodriguez Herrero, E.; Khodaparast, L.; Khodaparast, L.; Boon, N.; Bernaerts, K.; Quirynen, M.; Teughels, W. Development of antiseptic adaptation and cross-adapatation in selected oral pathogens in vitro. Sci. Rep. 2019, 9, 8326. [Google Scholar] [CrossRef] [PubMed]
- Bescos, R.; Ashworth, A.; Cutler, C.; Brookes, Z.L.; Belfield, L.; Rodiles, A.; Casas-Agustench, P.; Farnham, G.; Liddle, L.; Burleigh, M.; et al. Effects of Chlorhexidine mouthwash on the oral microbiome. Sci. Rep. 2020, 10, 5254. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.G.; Rosalen, P.L.; Falsetta, M.L.; Koo, H. Natural products in caries research: Current (limited) knowledge, challenges and future perspective. Caries Res. 2011, 45, 243–263. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.; Murata, R.M.; Duarte, S. Antimicrobial traits of tea- and cranberry-derived polyphenols against streptococcus mutans. Caries Res. 2011, 45, 327–335. [Google Scholar] [CrossRef]
- Cheng, L.; Li, J.; He, L.; Zhou, X. Natural products and caries prevention. Caries Res. 2015, 49, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Gazzani, G.; Daglia, M.; Papetti, A. Food components with anticaries activity. Curr. Opin. Biotechnol. 2012, 23, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Iha, K.; Suzuki, N.; Yoneda, M.; Takeshita, T.; Hirofuji, T. Effect of mouth cleaning with hinokitiol-containing gel on oral malodor: A randomized, open-label pilot study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2013, 116, 433–439. [Google Scholar] [CrossRef]
- Lodhia, P.; Yaegaki, K.; Khakbaznejad, A.; Imai, T.; Sato, T.; Tanaka, T.; Murata, T.; Kamoda, T. Effect of green tea on volatile sulfur compounds in mouth air. J. Nutr. Sci. Vitaminol. 2008, 54, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Toe, M.; Nagata, H.; Ojima, M.; Kuboniwa, M.; Shimizu, K.; Osawa, K.; Shizukuishi, S. Effect of Eucalyptus-Extract Chewing Gum on Oral Malodor: A Double-Masked, Randomized Trial. J. Periodontol. 2010, 81, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Gomes Muniz, F.W.M.; Nogueira, S.B.; Vasconcelos Mendes, F.L.; Rösing, C.K.; Mendes Moreira, M.M.S.; De Andrade, G.M.; De Sousa Carvalho, R. The impact of antioxidant agents complimentary to periodontal therapy on oxidative stress and periodontal outcomes: A systematic review. Arch. Oral Biol. 2015, 60, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; Davinelli, S.; Mangano, K.; Tempera, G.; Nicolosi, D.; Corsello, S.; Vergalito, F.; Tartaglia, E.; Scapagnini, G.; Di Marco, R. Effects of a new combination of plant extracts plus d-mannose for the management of uncomplicated recurrent urinary tract infections. J. Chemother. 2018, 30, 107–114. [Google Scholar] [CrossRef]
- Platel, K.; Srinivasan, K. Bioavailability of micronutrients from plant foods: An update. Crit. Rev. Food Sci. Nutr. 2016, 56, 1608–1619. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Liu, X.; Luo, Y.; Guo, H.; Hu, X.; Chen, F. Evaluation of Protective Effect of Freeze-Dried Strawberry, Grape, and Blueberry Powder on Acrylamide Toxicity in Mice. J. Food Sci. 2015, 80, H869–H874. [Google Scholar] [CrossRef] [PubMed]
- Kopec, R.E.; Cooperstone, J.L.; Schweiggert, R.M.; Young, G.S.; Harrison, E.H.; Francis, D.M.; Clinton, S.K.; Schwartz, S.J. Avocado consumption enhances human postprandial provitamin a absorption and conversion from a novel high-β-carotene tomato sauce and from carrots. J. Nutr. 2014, 144, 1158–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, H.M.; Shanmugaraj, B.M.; Srinivasan, B.; Ramalingam, S. Influence of Genotypic Variations on Antioxidant Properties in Different Fractions of Tomato. J. Food Sci. 2012, 77, C1174–C1178. [Google Scholar] [CrossRef] [PubMed]
- Bohlooli, S.; Barmaki, S.; Khoshkhahesh, F.; Nakhostin-Roohi, B. The effect of spinach supplementation on exercise-induced oxidative stress. J. Sports Med. Phys. Fit. 2015, 55, 609–614. [Google Scholar]
- Van Der Velden, U.; Kuzmanova, D.; Chapple, I.L.C. Micronutritional approaches to periodontal therapy. J. Clin. Periodontol. 2011, 38, 142–158. [Google Scholar] [CrossRef]
- Thompson, C.A.; Habermann, T.M.; Wang, A.H.; Vierkant, R.A.; Folsom, R.; Ross, J.A.; Cerhan, J.R. Antioxidant intake from fruits, vegetables and other sources and risk of non-Hodgkin’s lymphoma: The Iowa Women’s Health Study. Int. J. Cancer. 2010, 126, 992–1003. [Google Scholar] [CrossRef]
- Sdona, E.; Ekström, S.; Andersson, N.; Hallberg, J.; Rautiainen, S.; Håkansson, N.; Wolk, A.; Kull, I.; Melén, E.; Bergström, A. Fruit, vegetable and dietary antioxidant intake in school age, respiratory health up to young adulthood. Clin. Exp. Allergy 2021, 52, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ríos, P.; Pussinen, P.J.; Vernal, R.; Hernández, M. Oxidative stress in the local and systemic events of apical periodontitis. Front. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Quinn, P.J. The location and function of vitamin E in membranes (Review). Mol. Membr. Biol. 2000, 17, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.; Raza, S.T.; Ahmed, F.; Ahmad, A.; Abbas, S.; Mahdi, F. The role of Vitamin E in human health and some diseases. Sultan Qaboos Univ. Med. J. 2014, 14, 157–165. [Google Scholar]
- Lee, G.Y.; Han, S.N. The role of vitamin E in immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, U.; Devaraj, S.; Jialal, I. Vitamin E, oxidative stress, and inflammation. Annu. Rev. Nutr. 2005, 25, 151–174. [Google Scholar] [CrossRef]
- Ohta, Y.; Kobayashi, T.; Imai, Y.; Inui, K.; Yoshino, J.; Nakazawa, S. Effect of oral vitamin E administration on acute gastric mucosal lesion progression in rats treated with compound 48/80, a mast cell degranulator. Biol. Pharm. Bull. 2006, 29, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Kamsiah, J.; Muhaizan, W.; Gapor, M.; Roslin, O. Mucosal Protective Effects of Vitamin E on Aspirin-Induced Gastric Lesions in Rats. Int. J. Pharmacol. 2005, 1, 93–97. [Google Scholar]
- Kensche, A.; Reich, M.; Kümmerer, K.; Hannig, M.; Hannig, C. Lipids in preventive dentistry. Clin. Oral Investig. 2013, 17, 669–685. [Google Scholar] [CrossRef]
- Khader, M.; Eckl, P.M. Thymoquinone: An emerging natural drug with a wide range of medical applications. Iran. J. Basic Med. Sci. 2014, 17, 950–957. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Haas, B.; Grenier, D. Tea polyphenols inhibit the growth and virulence properties of Fusobacterium nucleatum. Sci. Rep. 2017, 7, 44815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gossweiler, A.G.; Martinez-Mier, E.A. Chapter 6: Vitamins and oral health. Monogr. Oral Sci. 2020, 28, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Beal, J.; Farny, N.G.; Haddock-Angelli, T.; Selvarajah, V.; Baldwin, G.S.; Buckley-Taylor, R.; Gershater, M.; Kiga, D.; Marken, J.; Sanchania, V.; et al. Robust estimation of bacterial cell count from optical density. Commun. Biol. 2020, 3, 512. [Google Scholar] [CrossRef] [PubMed]
- McBirney, S.E.; Trinh, K.; Wong-Beringer, A.; Armani, A.M. Wavelength-normalized spectroscopic analysis of Staphylococcus aureus and Pseudomonas aeruginosa growth rates. Biomed. Opt. Express 2016, 7, 4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Stepanović, S.; Ćirković, I.; Ranin, L.; Švabić-Vlahović, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef]
- Wilson, C.; Rutering, J.; Ilmer, M.; Recio, A.; Coleman, M.; Vykoukal, J.; Alt, E.; Orleans, N. Quantitative and Qualitative Assessment Methods for Biofilm Growth: A Mini-review Christina. Nat. Rev. Drug Discov. 2016, 5, 1–8. [Google Scholar]
- Mahon, C.R.; Lehman, D.C.; Manuselis, G. Textbook of Diagnostic Microbiology-E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Pommerville, J.C. Alcamo’s Fundamentals of Microbiology; Jones and Bartlett Publishers: Burlington, MA, USA, 2004. [Google Scholar]
- Anderson, A.C.; Rothballer, M.; Altenburger, M.J.; Woelber, J.P.; Karygianni, L.; Lagkouvardos, I.; Hellwig, E.; Al-Ahmad, A. In-vivo shift of the microbiota in oral biofilm in response to frequent sucrose consumption. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen NIH Public Access. Curr. Opin. Microbiol. 2016, 23, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basic, A.; Blomqvist, M.; Dahlén, G.; Svensäter, G. The proteins of Fusobacterium spp. involved in hydrogen sulfide production from L-cysteine. BMC Microbiol. 2017, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bidossi, A.; Bortolin, M.; Toscano, M.; De Vecchi, E.; Romanò, C.L.; Mattina, R.; Drago, L. In vitro comparison between α-tocopheryl acetate and α-tocopheryl phosphate against bacteria responsible of prosthetic and joint infections. PLoS ONE 2017, 12, e1–e12. [Google Scholar] [CrossRef] [Green Version]
- Vergalito, F.; Pietrangelo, L.; Petronio Petronio, G.; Colitto, F.; Alfio Cutuli, M.; Magnifico, I.; Venditti, N.; Guerra, G.; Di Marco, R. Vitamin E for prevention of biofilm-caused Healthcare-associated infections. Open Med. 2020, 15, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Camargo Smolarek, P.; Esmerino, L.A.; Chibinski, A.C.; Bortoluzzi, M.C.; dos Santos, E.B.; Kozlowski, V.A. In vitro antimicrobial evaluation of toothpastes with natural compounds. Eur. J. Dent. 2015, 9, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Thorgeirsdóttir, T.Ó.; Kristmundsdóttir, T.; Thormar, H.; Axelsdóttir, Í.; Holbrook, W.P. Antimicrobial activity of monocaprin: A monoglyceride with potential use as a denture disinfectant. Acta Odontol. Scand. 2006, 64, 21–26. [Google Scholar] [CrossRef]
- Kim, S.A.; Rhee, M.S. Highly enhanced bactericidal effects of medium chain fatty acids (caprylic, capric, and lauric acid) combined with edible plant essential oils (carvacrol, eugenol, β-resorcylic acid, trans-cinnamaldehyde, thymol, and vanillin) against Escherichia coli O15. Food Control 2016, 60, 447–454. [Google Scholar] [CrossRef]
- Cho, T.J.; Park, S.M.; Yu, H.; Seo, G.H.; Kim, H.W.; Kim, S.A.; Rhee, M.S. Recent advances in the application of antibacterial complexes using essential oils. Molecules 2020, 25, 1752. [Google Scholar] [CrossRef] [PubMed]
- Chapple, I.L.; Matthews, J.B.; Wright, H.J.; Scott, A.E.; Griffiths, H.R.; Grant, M.M. Ascorbate and α-tocopherol differentially modulate reactive oxygen species generation by neutrophils in response to FcγR and TLR agonists. Innate Immun. 2013, 19, 152–159. [Google Scholar] [CrossRef]
- Liu, J.; Ling, J.; Wu, C.D. ScienceDirect Cetylpyridinium chloride suppresses gene expression associated with halitosis. Arch. Oral Biol. 2013, 8, 2–7. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Active Compounds | Effect on Oral Microbiota/Tissues | References |
|---|---|---|---|
| Tea, propolis, cranberry, Galla chinensis, grapes, coffee, cacao | Polyphenols | Reduction of oral biofilm | [20,25,26,27,28] |
| Hinokitiol (from Cupressaceae trees), green tea powder, eucalyptus extract, oil from N. sativa seeds | Terpenoids, terpenes, theaflavins, catechins | Reduction of oral malodor | [19,29,30,31,50,51] |
| Blueberries, strawberries, grapes, avocado, tomatoes, spinach, and carrots | Fat-soluble vitamins (vitamin A, vitamin E-tocopherol and b-carotene); water-soluble vitamins (vitamin C and vitamin B complex); trace elements (zinc, magnesium); and bioflavonoids (plant derived) | Oxidative stress was demonstrated as being causative of oral dysbiosis and oral pathologies. Antioxidant compounds counteract the oxidative stress factors restoring oral eubiosis | [4,32,33,34,35,36,37,38,39,42] |
| Oils from seeds (almonds, sunflowers, pine nuts, olives, peanuts, coconut, corn, hemp, wheat, etc.) | Vitamin E, alpha-tocopherol | Treatment with vitamin E induces in the oral cavity a sensitive decrease in plaque and biofilm formation, probing depth, clinical attachment level, and bleeding on probing | [39,42,49,52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrangelo, L.; Magnifico, I.; Petronio Petronio, G.; Cutuli, M.A.; Venditti, N.; Nicolosi, D.; Perna, A.; Guerra, G.; Di Marco, R. A Potential “Vitaminic Strategy” against Caries and Halitosis. Appl. Sci. 2022, 12, 2457. https://doi.org/10.3390/app12052457
Pietrangelo L, Magnifico I, Petronio Petronio G, Cutuli MA, Venditti N, Nicolosi D, Perna A, Guerra G, Di Marco R. A Potential “Vitaminic Strategy” against Caries and Halitosis. Applied Sciences. 2022; 12(5):2457. https://doi.org/10.3390/app12052457
Chicago/Turabian StylePietrangelo, Laura, Irene Magnifico, Giulio Petronio Petronio, Marco Alfio Cutuli, Noemi Venditti, Daria Nicolosi, Angelica Perna, Germano Guerra, and Roberto Di Marco. 2022. "A Potential “Vitaminic Strategy” against Caries and Halitosis" Applied Sciences 12, no. 5: 2457. https://doi.org/10.3390/app12052457
APA StylePietrangelo, L., Magnifico, I., Petronio Petronio, G., Cutuli, M. A., Venditti, N., Nicolosi, D., Perna, A., Guerra, G., & Di Marco, R. (2022). A Potential “Vitaminic Strategy” against Caries and Halitosis. Applied Sciences, 12(5), 2457. https://doi.org/10.3390/app12052457