
The Antioxidant Effect of Natural Antimicrobials in Shrimp Primary Intestinal Cells Infected with Nematopsis messor

 ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasites and Antimicrobials
2.2. In Vitro Infection Assay in Primary Shrimp Gut Epithelial Cells (SGP)
2.3. Gene Expression Analysis
2.4. Transepithelial Resistance of Cellular Tight Junctions (TEER) and Actin Polymerization Assay
2.5. Challenge Tests (Counting Living Larvae)
2.6. H2O2 Production in SGP Cells following Infection
2.7. Determination of Natural Antimicrobial LC50 Cytotoxicity and the Impact on Epithelial Cell Proliferation (MTT Assay)
2.8. Measurement of SOD and CAT Activity in SGP Cells and Gut Tissue
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
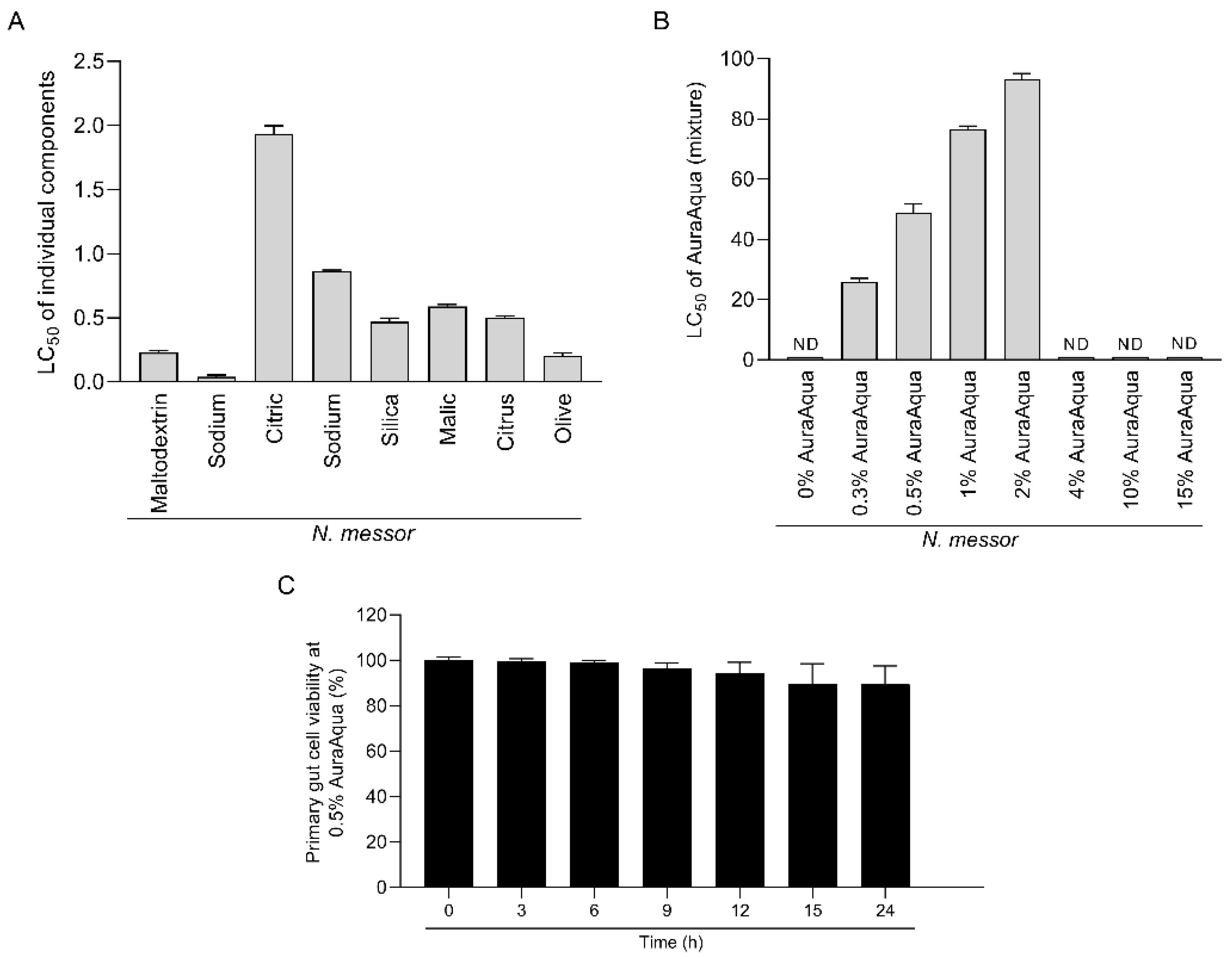
3.1. In Vitro Cytotoxic Effects of the Antimicrobial Mixture on SGP Cells
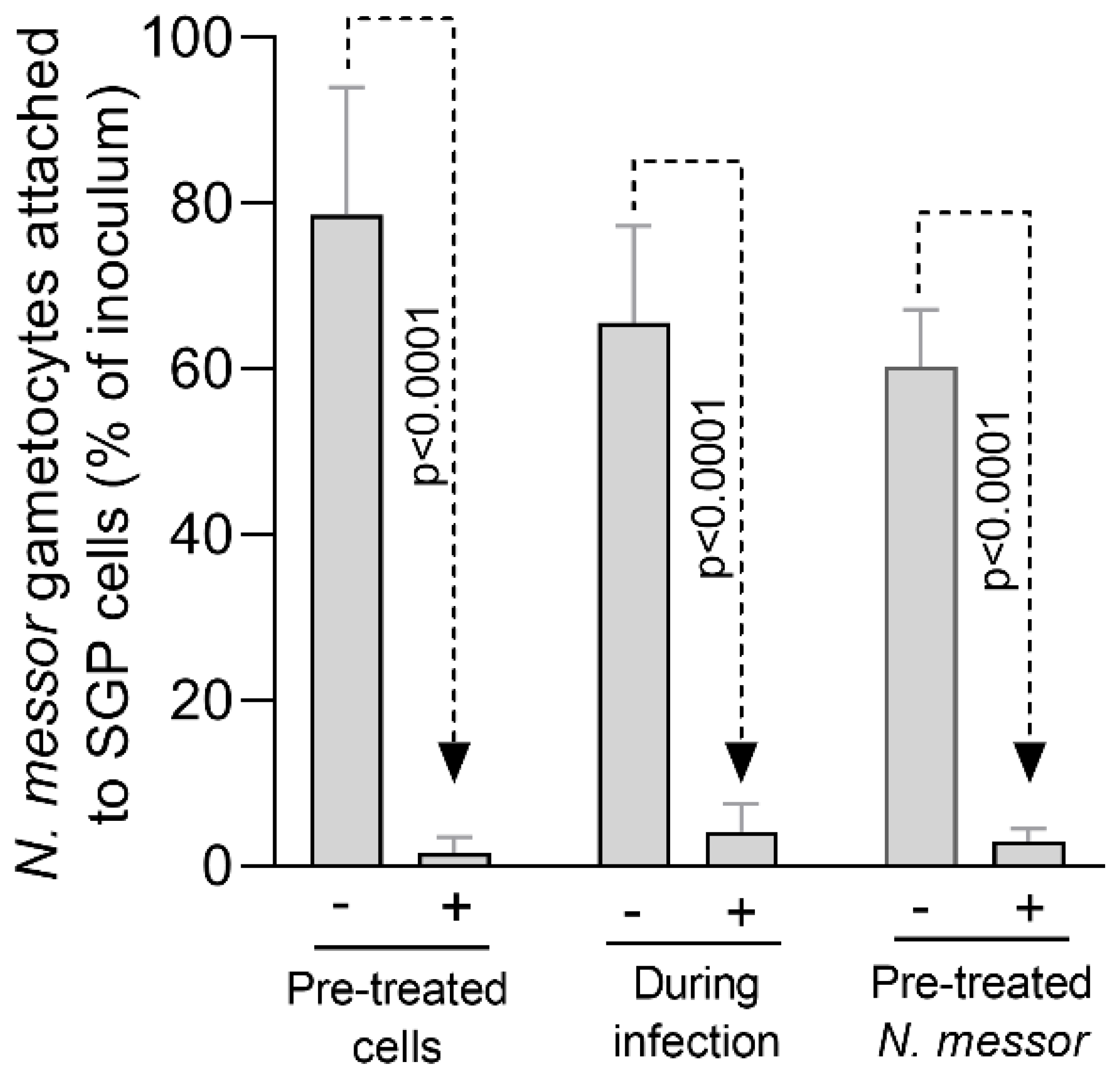
3.2. In Vitro Effect of Aq on N. messor Infection of SGP Cells
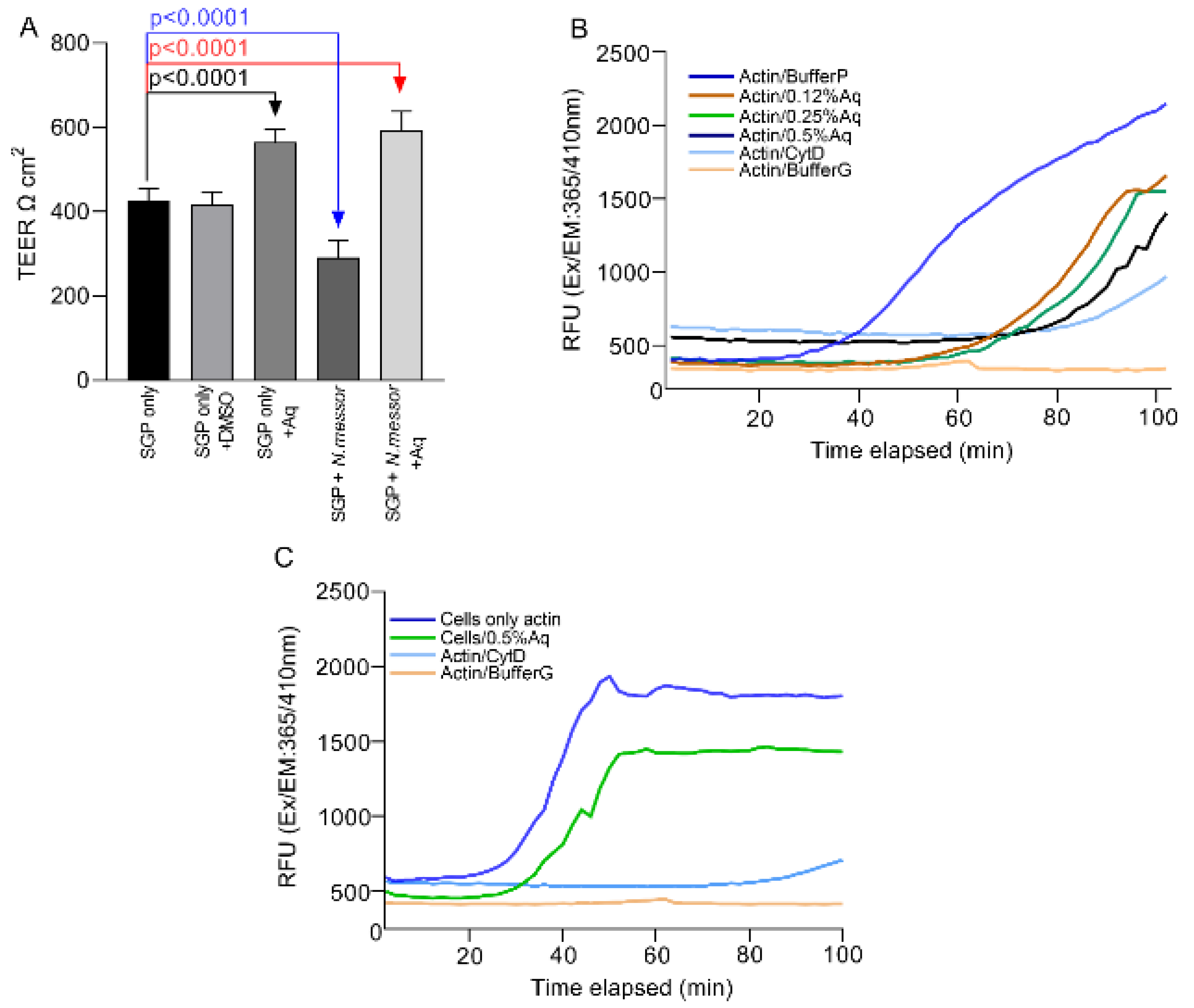
3.3. Natural Antimicrobials Strengthens Cellular Tight Junctions (TEER) and Prevents Actin Polymerisation during Infection
3.4. The Impact of Aq on the Induced Oxidative Stress in N. messor Infected SGP Cells
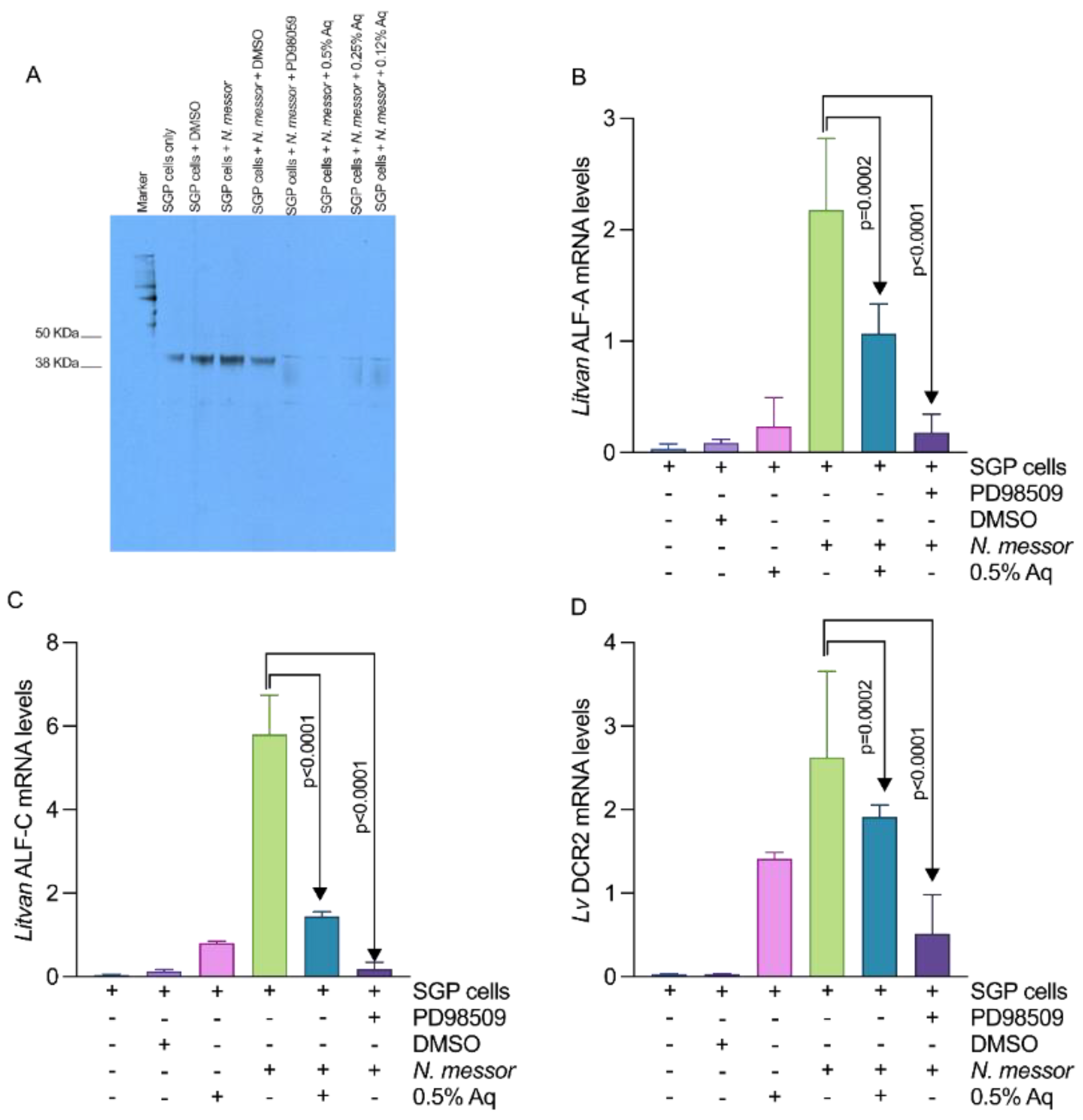
3.5. AuraAqua Inhibits the Production of Immune-Related Antimicrobial Peptides in Infected SGP Cells by Blocking the ERK-Signaling Pathway
3.6. Challenge Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seeber, F.; Steinfelder, S. Recent advances in understanding apicomplexan parasites. F1000Research 2016, 5, 1369. [Google Scholar] [CrossRef] [PubMed]
- Prasadan, K.P.; Janardanan, K.P. Three New Species of Gregarines (Apicomplexa: Sporozoea: Porosporidae) in the Estu-arine Crabs from Kerala, India. Acta Protozoologica 2001, 40, 303–309. [Google Scholar]
- Jiménez, R.; De Barniol, L.; Machuca, M. Nematopsis marinus n. sp., a new septate gregarine from cultured penaeoid shrimp Litopenaeus vannamei (Boone), in Ecuador. Aquac. Res. 2002, 33, 231–240. [Google Scholar] [CrossRef]
- Lightner, D.V. A Handbook of Shrimp Pathology and Diagnostic Procedures for Diseases of Cultured Penaeid Shrimp; World Aquaculture Society: Baton Rouge, LA, USA, 1996. [Google Scholar]
- Ibrahim, N.; Ibrahim, H.; Sabater, A.M.; Mazier, D.; Valentin, A.; Nepveu, F. Artemisinin nanoformulation suitable for intravenous injection: Preparation, characterization and antimalarial activities. Int. J. Pharm. 2015, 495, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Sima, F.; Stratakos, A.C.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Gundogdu, O.; Lazar, V.; Corcionivoschi, N. A Novel Natural Antimicrobial Can Reduce the in vitro and in vivo Pathogenicity of T6SS Positive Campylobacter jejuni and Campylobacter coli Chicken Isolates. Front. Microbiol. 2018, 9, 2139. [Google Scholar] [CrossRef] [PubMed]
- Centrone, M.; D’Agostino, M.; Difonzo, G.; De Bruno, A.; Di Mise, A.; Ranieri, M.; Montemurro, C.; Valenti, G.; Poiana, M.; Caponio, F.; et al. Antioxidant Efficacy of Olive By-Product Extracts in Human Colon HCT8 Cells. Foods 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Ashour, D.S. Toll-like receptor signaling in parasitic infections. Expert Rev. Clin. Immunol. 2015, 11, 771–780. [Google Scholar] [CrossRef]
- Buret, A.G.; Mitchell, K.; Muench, D.G.; Scott, K.G.E. Giardia lamblia disrupts tight junctional ZO-1 and increases permeability in non-transformed human small intestinal epithelial monolayers: Effects of epidermal growth factor. Parasitology 2002, 125, 11–19. [Google Scholar] [CrossRef]
- Jones, J.E.; Korcsmaros, T.; Carding, S.R. Mechanisms and pathways of Toxoplasma gondii transepithelial migration. Tissue Barriers 2017, 5, e1273865. [Google Scholar] [CrossRef]
- Da Costa, R.F.; Souza, W.D.; Benchimol, M.; Alderete, J.F.; Morgado-Diaz, J.A. Trichomonas vaginalis perturbs the junctional complex in epithelial cells. Cell Res. 2005, 15, 704–716. [Google Scholar] [CrossRef]
- Szewczyk-Golec, K.; Pawłowska, M.; Wesołowski, R.; Wróblewski, M.; Mila-Kierzenkowska, C. Oxidative Stress as a Possible Target in the Treatment of Toxoplasmosis: Perspectives and Ambi-guities. Int. J. Mol. Sci. 2021, 22, 5705. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bandyapadhyay, R.; Basu, M.K. Leishmania donovani: Superoxide dismutase level in infected macrophages. Biosci. Rep. 1988, 8, 131–137. [Google Scholar] [CrossRef] [PubMed]
- McCall, R.M.; Frei, B. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic. Biol. Med. 1999, 26, 1034–1053. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, Y.; García-Heredia, A.; Merino-Mascorro, A.; García, S.; Solís-Soto, L.; Heredia, N. Natural and synthetic antimicrobials reduce adherence of enteroaggregative and enterohemorrhagic Escherichia coli to epithelial cells. PLoS ONE 2021, 16, e0251096. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Stef, L.; Pet, I.; Ward, P.; Callaway, T.; Ricke, S.C.; Gundogdu, O.; Corcionivoschi, N. Antiviral activity of a novel mixture of natural antimicrobials, in vitro, and in a chicken infection model in vivo. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Schink, A.; Neumann, J.; Leifke, A.L.; Ziegler, K.; Fröhlich-Nowoisky, J.; Cremer, C.; Thines, E.; Weber, B.; Pöschl, U.; Schuppan, D.; et al. Screening of herbal extracts for TLR2- and TLR4-dependent anti-inflammatory effects. PLoS ONE 2018, 13, e0203907. [Google Scholar] [CrossRef]
- Rastew, E.; Vicente, J.; Singh, U. Oxidative stress resistance genes contribute to the pathogenic potential of the anaerobic protozoan parasite, Entamoeba histolytica. Int. J. Parasitol. 2012, 42, 1007–1015. [Google Scholar] [CrossRef]
- Guyton, K.Z.; Liu, Y.; Gorospe, M.; Xu, Q.; Holbrook, N.J. Activation of Mitogen-activated Protein Kinase by H2O2. J. Biol. Chem. 1996, 271, 4138–4142. [Google Scholar] [CrossRef]
- Munoz-Caro, T.; Mena Huertas, S.J.; Conejeros, I.; Alarcon, P.; Hidalgo, M.A.; Burgos, R.A.; Hermosilla, C.; Taubert, A. Eimeria bovis-triggered neutrophil extracellular trap formation is CD11b-, ERK 1/2-, p38 MAP kinase- and SOCE-dependent. Vet. Res. 2015, 46, 23. [Google Scholar] [CrossRef]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Et Biophys. Acta 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Y.; Li, C.; Ai, K.; Li, K.; Li, H.; Yang, J. The evolutionarily conserved MAPK/Erk signaling promotes ancestral T-cell immunity in fish via c-Myc–mediated glycolysis. J. Biol. Chem. 2020, 295, 3000–3016. [Google Scholar] [CrossRef] [PubMed]
- Helfenberger, K.E.; Villalba, N.M.; Buchholz, B.; Boveris, A.; Poderoso, J.J.; Gelpi, R.J.; Poderoso, C. Subcellular distribution of ERK phosphorylation in tyrosine and threonine depends on redox status in murine lung cells. PLoS ONE 2018, 13, e0193022. [Google Scholar] [CrossRef] [PubMed]
- Lorgeril, D.; Julien; Gueguen, Y.; Goarant, C.; Goyard, E.; Mugnier, C.; Fievet, J.; Piquemal, D.; Bachère, E. A relationship between antimicrobial peptide gene expression and capacity of a selected shrimp line to survive a Vibrio infection. Mol. Immunol. 2008, 45, 3438–3445. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, H.; Li, S.; Li, F.; Xiang, J. Structure and Bioactivity of a Modified Peptide Derived from the LPS-Binding Domain of an An-ti-Lipopolysaccharide Factor (ALF) of Shrimp. Mar. Drugs 2016, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Supungul, P.; Klinbunga, S.; Pichyangkura, R.; Jitrapakdee, S.; Hirono, I.; Aoki, T.; Tassanakajon, A. Identification of immune-related genes in hemocytes of black tiger shrimp (Penaeus monodon). Mar. Biotechnol. 2002, 4, 487–494. [Google Scholar] [CrossRef]
- Silveira, A.S.; Matos, G.M.; Falchetti, M.; Ribeiro, F.S.; Bressan, A.; Bachère, E.; Perazzolo, L.M.; Rosa, R.D. An immune-related gene expression atlas of the shrimp digestive system in response to two major pathogens brings insights into the involvement of hemocytes in gut immunity. Dev. Comp. Immunol. 2018, 79, 44–50. [Google Scholar] [CrossRef]
- Van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; van Houdt, W.; van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef]
- Castellanos-Gonzalez, A.; Cabada, M.M.; Nichols, J.; Gomez, G.; White, A.C. Human Primary Intestinal Epithelial Cells as an Improved In Vitro Model for Cryptosporidium parvum Infection. Infect. Immun. 2013, 81, 1996–2001. [Google Scholar] [CrossRef]
- Jayesh, P.; Seena, J.; Singh, I.S.B. Establishment of shrimp cell lines: Perception and orientation. Indian J. Virol. Off. Organ Indian Virol. Soc. 2012, 23, 244–251. [Google Scholar] [CrossRef]
- Peh, E.; Kittler, S.; Reich, F.; Kehrenberg, C. Antimicrobial activity of organic acids against Campylobacter spp. and development of combinations—A synergistic effect? PLoS ONE 2020, 15, e0239312. [Google Scholar] [CrossRef] [PubMed]
- Subash, P.; Uma, A.; Ahilan, B. Early responses in Penaeus vannamei during experimental infection with Enterocytozoon hepatopenaei (EHP) spores by injection and oral routes. J. Invertebr. Pathol. 2022, 190, 107740. [Google Scholar] [CrossRef] [PubMed]
- Checkley, W.; White, A.C.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.-M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect. Dis. 2014, 15, 85–94. [Google Scholar] [CrossRef]
- Chakraborti, J.; Bandyopadhyay, P.K. First record of a parasitic septate gregarines (Apicomplexa: Sporozoea) in the shrimp Peneaus monodon in Sundarbans of West Bengal. J. Parasit. Dis. 2010, 34, 40–43. [Google Scholar] [CrossRef][Green Version]
- Balta, I.; Marcu, A.; Linton, M.; Kelly, C.; Gundogdu, O.; Stef, L.; Pet, I.; Ward, P.; Deshaies, M.; Callaway, T.; et al. Mixtures of natural antimicrobials can reduce Campylobacter jejuni, Salmonella enterica and Clostridium perfringens infections and cellular inflammatory response in MDCK cells. Gut Pathog. 2021, 13, 1–13. [Google Scholar] [CrossRef]
- Cardona, E.; Saulnier, D.; Lorgeoux, B.; Chim, L.; Gueguen, Y. Rearing effect of biofloc on antioxidant and antimicrobial transcriptional response in Litopenaeus stylirostris shrimp facing an experimental sub-lethal hydrogen peroxide stress. Fish Shellfish Immunol. 2015, 45, 933–939. [Google Scholar] [CrossRef]
- Rosa, R.D.; Vergnes, A.; De Lorgeril, J.; Goncalves, P.; Perazzolo, L.M.; Saune, L.; Romestand, B.; Fievet, J.; Gueguen, Y.; Bachere, E.; et al. Functional divergence in shrimp anti-lipopolysaccharide factors (ALFs): From recognition of cell wall com-ponents to antimicrobial activity. PLoS ONE 2013, 8, e67937. [Google Scholar] [CrossRef]
- Chen, Y.H.; Jia, X.T.; Zhao, L.; Li, C.Z.; Zhang, S.; Chen, Y.G.; Weng, S.P.; He, J.G. Identification and functional characterization of Dicer2 and five single VWC domain proteins of Litopenaeus vannamei. Dev. Comp. Immunol. 2011, 35, 661–671. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Ranieri, M.; DI Mise, A.; Difonzo, G.; Centrone, M.; Venneri, M.; Pellegrino, T.; Russo, A.; Mastrodonato, M.; Caponio, F.; Valenti, G.; et al. Green olive leaf extract (OLE) provides cytoprotection in renal cells exposed to low doses of cadmium. PLoS ONE 2019, 14, e0214159. [Google Scholar] [CrossRef]
- Sotomayor, M.A.; Reyes, J.K.; Restrepo, L.; Domínguez-Borbor, C.; Maldonado, M.; Bayot, B. Efficacy assessment of commercially available natural products and antibiotics, commonly used for mitigation of pathogenic Vibrio outbreaks in Ecuadorian Penaeus (Litopenaeus) vannamei hatcheries. PLoS ONE 2019, 14, e0210478. [Google Scholar] [CrossRef] [PubMed]
- Corcionivoschi, N.; Alvarez, L.A.; Sharp, T.H.; Strengert, M.; Alemka, A.; Mantell, J.; Verkade, P.; Knaus, U.G.; Bourke, B. Mucosal reactive oxygen species decrease virulence by disrupting Campylobacter jejuni phos-photyrosine signaling. Cell Host Microbe 2012, 12, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Remmal, A.; Achahbar, S.; Bouddine, L.; Chami, N.; Chami, F. In vitro destruction of Eimeria oocysts by essential oils. Veter. Parasitol. 2011, 182, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lu, M.; Zhang, C.; Wu, X.; Chen, J.; Lv, W.; Sun, T.; Qiu, H.; Huang, S. Oxidative stress modulates the expression of toll-like receptor 3 during respiratory syncytial virus infection in human lung epithelial A549 cells. Mol. Med. Rep. 2018, 18, 1867–1877. [Google Scholar] [CrossRef]
- Ardiansyah; Jaya, A.A.; Amrullah; Dahlia; Baiduri, M.A.; Hartinah; Wahidah; Indrayani. Antioxidant status and oxidative stress markers of white faeces syndrome-infected Pacific white shrimp (Litopenaeus vannamei Boone). Aacl. Bioflux 2020, 13, 503–517. [Google Scholar]
- Chirawithayaboon, P. Hepatopancreatic antioxidant enzyme activities and disease resistance of Pacific white shrimp (Li-topenaeus vannamei) fed diet supplemented with garlic (Allium sativum) extract. Agric. Nat. Resour. 2020, 54, 377–386. [Google Scholar]
- Johny, S.; Merisko, A.; Whitman, D.W. Efficacy of eleven antimicrobials against a gregarine parasite (Apicomplexa: Pro-tozoa). Ann. Clin. Microbiol. Antimicrob. 2007, 6, 15. [Google Scholar] [CrossRef]
- Fajer-Ávila, E.J.; Covarrubias, M.S.M.; Abad-Rosales, S.; Roque, A.; Meza-Bojórquez, P.; Hernández-González, C. Effectiveness of oral Elancoban™ and Avimix-ST™ against Nematopsis (Apicomplexa: Porosporidae) gametocyts infecting the shrimp Litopenaeus vannamei. Aquaculture 2005, 244, 11–18. [Google Scholar] [CrossRef]
- Balta, I.; Marcu, A.; Linton, M.; Kelly, C.; Stef, L.; Pet, I.; Ward, P.; Pircalabioru, G.G.; Chifiriuc, C.; Gundogdu, O.; et al. The in vitro and in vivo anti-virulent effect of organic acid mixtures against Eimeria tenella and Eimeria bovis. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Revoltella, S.; Baraldo, G.; Waltenberger, B.; Schwaiger, S.; Kofler, P.; Moesslacher, J.; Huber-Seidel, A.; Pagitz, K.; Kohl, R.; Jansen-Duerr, P.; et al. Identification of the NADPH Oxidase 4 Inhibiting Principle of Lycopus europaeus. Molecules 2018, 23, 653. [Google Scholar] [CrossRef]
- Gardner, R.; Salvador, A.; Moradas-Ferreira, P. Why does SOD overexpression sometimes enhance, sometimes decrease, hydrogen peroxide production? a minimalist explanation. Free Radic. Biol. Med. 2002, 32, 1351–1357. [Google Scholar] [CrossRef]
- Hashim, A.; Mulcahy, G.; Bourke, B.; Clyne, M. Interaction of Cryptosporidium hominis and Cryptosporidium parvum with Primary Human and Bovine Intestinal Cells. Infect. Immun. 2006, 74, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Ch Stratakos, A.; Sima, F.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Pet, I.; Iancu, T.; Pircalabioru, G.; et al. The in vitro and ex vivo effect of Auranta 3001 in preventing Cryptosporidium hominis and Cryptosporidium parvum infection. Gut Pathog 2017, 9, 49. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goddette, D.W.; Frieden, C. Actin polymerization. The mechanism of action of cytochalasin D. J. Biol. Chem. 1986, 261, 15974–15980. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Tonelli, R.R. Infection Strategies of Intestinal Parasite Pathogens and Host Cell Responses. Front. Microbiol. 2016, 7, 256. [Google Scholar] [CrossRef] [PubMed]
- Lucera, A.; Costa, C.; Conte, A.; Del Nobile, M.A. Food applications of natural antimicrobial compounds. Front. Microbiol. 2012, 3, 287. [Google Scholar] [CrossRef] [PubMed]
- Kraft, M.R.; Klotz, C.; Bücker, R.; Schulzke, J.D.; Aebischer, T. Giardia’s Epithelial Cell Interaction In Vitro: Mimicking Asymptomatic Infection? Front. Cell. Infect. Microbiol. 2017, 7, 421. [Google Scholar] [CrossRef]
- Matos, G.M.; Schmitt, P.; Barreto, C.; Farias, N.D.; Toledo-Silva, G.; Guzmán, F.; Destoumieux-Garzón, D.; Perazzolo, L.M.; Rosa, R.D. Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp. Mar. Drugs 2018, 16, 381. [Google Scholar] [CrossRef]
- Johansson, M.; Lind, M.; Holmblad, T.; Thornqvist, P.; Soderhall, K. Peroxinectin, a Novel Cell Adhesion Protein from Crayfish Blood. Biochem. Biophys. Res. Commun. 1995, 216, 1079–1087. [Google Scholar] [CrossRef]
- Dong, C.; Wei, Z.; Yang, G. Involvement of peroxinectin in the defence of red swamp crayfish Procambarus clarkii against pathogenic Aeromonas hydrophila. Fish Shellfish Immunol. 2011, 30, 1223–1229. [Google Scholar] [CrossRef]
- Sritunyalucksana, K.; Wongsuebsantati, K.; Johansson, M.; Söderhäll, K. Peroxinectin, a cell adhesive protein associated with the proPO system from the black tiger shrimp, Penaeus monodon. Dev. Comp. Immunol. 2001, 25, 353–363. [Google Scholar] [CrossRef]
- Jin, X.; Gong, P.; Zhang, X.; Li, G.; Zhu, T.; Zhang, M.; Li, J. Activation of ERK Signaling via TLR11 Induces IL-12p40 Production in Peritoneal Macrophages Challenged by Neospora caninum. Front. Microbiol. 2017, 8, 1393. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, J.; Lin, L.; Pan, G.; Zhang, S.; Chen, H.; Zhang, M.; Xuan, Y.; Wang, Y.; You, Z. Quzhou Fructus Aurantii Extract suppresses inflammation via regulation of MAPK, NF-κB, and AMPK signaling pathway. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chei, S.; Oh, H.-J.; Song, J.-H.; Seo, Y.-J.; Lee, K.; Kim, K.-J.; Lee, B.-Y. Spirulina maxima extract prevents activation of the NLRP3 inflammasome by inhibiting ERK signaling. Sci. Rep. 2020, 10, 2075. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.K.; Koh, C.B.; Teoh, C.Y.; Romano, N. Farm-raised tiger shrimp, Penaeus monodon, fed commercial feeds with added organic acids showed en-hanced nutrient utilization, immune response and resistance to Vibrio harveyi challenge. Aquaculture 2015, 449, 69–77. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Accession Numbers | Primer Sequence Forward/Reverse (5′-3′) | Reference |
|---|---|---|---|
| SOD | XM_027376215.1 | (F) GCAATGAATGCCCTTCTACC(R) CAGAGCCTTTCACTCCAACG | [37] |
| Litvan ALF-A | EW713395 | (F) CTGATTGCTCTTGTGCCACG(R) TGACCCATGAACTCCACCTC | [38] |
| Litvan ALF-C | FE058235 | (F) ATGCGAGTGTCTGTCCTCAG(R) TGAGTTTGTTCGCGATGGCC | [38] |
| Lv DCR2 | HQ541163 | (F) AGGAAATGCAATGTCGTGGTT(R) ACGAGCCCTCCCCCTAGATT | [39] |
| Zo-1 | NM_0013011025.3 | (F) CGGGACTGTTGGTATTGGCTAGA(R) GGCCAGGGCCATAGTAAAGTTTG. | [36] |
| Occludin | NM_001205254.2 | (F) TCCTATAAATCCACGCCGGTTC(R) CTCAAAGTTACCACCGCTGCTG. | [36] |
| Mortality | Aq (%) | |
| 0 | 0.5 | |
| 68 ± 0.45 | 14 ± 1.22 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balta, I.; Stef, L.; Butucel, E.; Gradisteanu Pircalabioru, G.; Venig, A.; Ward, P.; Deshaies, M.; Pet, I.; Stef, D.; Koyun, O.Y.; et al. The Antioxidant Effect of Natural Antimicrobials in Shrimp Primary Intestinal Cells Infected with Nematopsis messor. Antioxidants 2022, 11, 974. https://doi.org/10.3390/antiox11050974
Balta I, Stef L, Butucel E, Gradisteanu Pircalabioru G, Venig A, Ward P, Deshaies M, Pet I, Stef D, Koyun OY, et al. The Antioxidant Effect of Natural Antimicrobials in Shrimp Primary Intestinal Cells Infected with Nematopsis messor. Antioxidants. 2022; 11(5):974. https://doi.org/10.3390/antiox11050974
Chicago/Turabian StyleBalta, Igori, Lavinia Stef, Eugenia Butucel, Gratiela Gradisteanu Pircalabioru, Adelina Venig, Patrick Ward, Myriam Deshaies, Ioan Pet, Ducu Stef, Osman Y. Koyun, and et al. 2022. "The Antioxidant Effect of Natural Antimicrobials in Shrimp Primary Intestinal Cells Infected with Nematopsis messor" Antioxidants 11, no. 5: 974. https://doi.org/10.3390/antiox11050974
APA StyleBalta, I., Stef, L., Butucel, E., Gradisteanu Pircalabioru, G., Venig, A., Ward, P., Deshaies, M., Pet, I., Stef, D., Koyun, O. Y., Callaway, T. R., Gundogdu, O., & Corcionivoschi, N. (2022). The Antioxidant Effect of Natural Antimicrobials in Shrimp Primary Intestinal Cells Infected with Nematopsis messor. Antioxidants, 11(5), 974. https://doi.org/10.3390/antiox11050974