Critical Impact of Different Conserved Endoplasmic Retention Motifs and Dopamine Receptor Interacting Proteins (DRIPs) on Intracellular Localization and Trafficking of the D2 Dopamine Receptor (D2-R) Isoforms

 , , , ,
, , , ,
Abstract
:1. Dopamine Receptors
2. Dopamine Receptor Type 2 (D2-R)
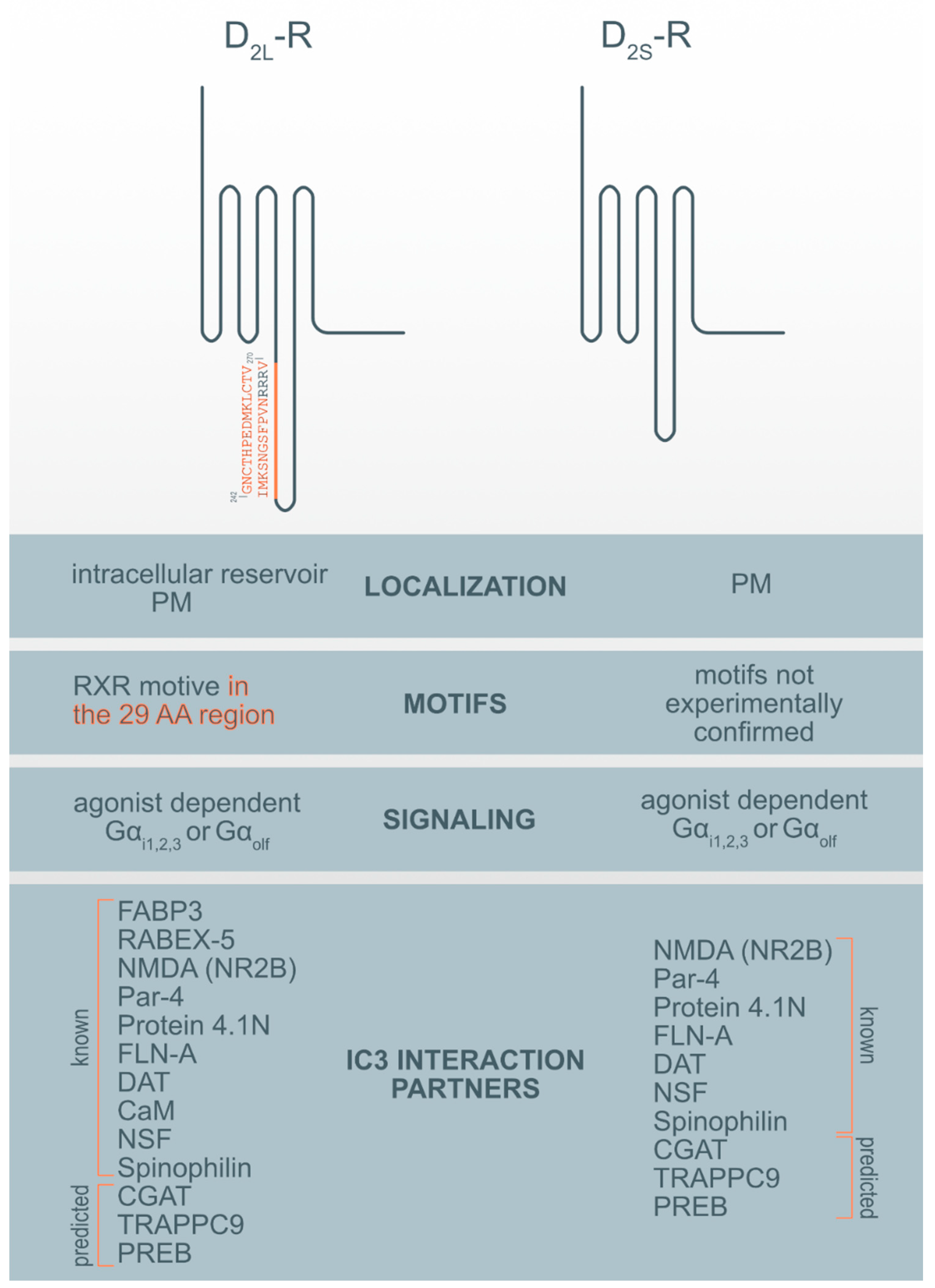
3. Localization Differences Between D2S-R and D2L-R
4. Functional Differences between D2S-R and D2L-R
5. The ER Retention Motifs in GPCRs and Both D2-R Isoforms
6. KDEL and KKXX Motifs
7. RXR and RSRR Motifs
8. ER Export Motifs in GPCRs and D2-R
9. D2-R Interaction Proteins (DRIPs)

{kind=link}
| Interaction Partner | S/N Ratio | Function | Reference |
|---|---|---|---|
| Glutamate, NMDA (NR2B) | 62.39 | ionotropic glutamate receptor | Liu, X.Y. et al. (2006) [82] |
| Par-4 | 48.63 | regulatory component in dopamine signaling | Guo, Q. et al. (1998) [83] Park, S.K. et al. (2005) [84] |
| Protein 4.1N | 38.61 | membrane-cytoskeleton adaptor | Binda, A.V. et al. (2002) [85] |
| FLN-A | 26.65 | actin binding protein | Lin, R. et al. (2001) [86] |
| DAT | 20.29 | facilitating reuptake of extracellular dopamine back in the cytosol | Lee, F.J. et al. (2007) [87] |
| Gα i/z/o | 17.85 | binding GPCRs | |
| CaM | 13.36 | intermediate calcium-binding messenger | Navarro, G. et al. (2009) [88] |
| NSF | 13.03 | ATPase | Hanson, P.I. et. al. (1995) [90] Zou S. et al. (2005) [91] |
| Spinophilin | 12.14 | F-actin and protein phosphatase-1-binding protein | Smith, F.D. et al. (1999) [92] |
| Predicted Interaction Partner | 12.14 | ||
| CGAT | 19.90 | involved in the transport of biogenic monoamines | |
| TRAPPC9 | 19.73 | involved in vesicular trafficking from ER to GA | |
| PREB | 18.78 | transcriptional regulator |
10. Interaction with ER-Resident Gatekeeper Proteins
11. Clinical Relevance
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jackson, D.M.; Westlind-Danielsson, A. Dopamine receptors: Molecular biology, biochemistry and behavioural aspects. Pharmacol. Ther. 1994, 64, 291–370. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Espinoza, S.; Gainetdinov, R.R. Dopamine receptors—IUPHAR Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, C.; Zheng, M.; Zhang, X.; Guo, S.; Kwon, K.J.; Shin, C.Y.; Kim, H.S.; Cheon, S.H.; Kim, K.M. N-linked Glycosylation on the N-terminus of the dopamine D2 and D3 receptors determines receptor association with specific microdomains in the plasma membrane. Biochim. Biophys. Acta 2015, 1853, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivonello, R.; Ferone, D.; Lombardi, G.; Colao, A.; Lamberts, S.W.J.; Hofland, L.J. Novel insights in dopamine receptor physiology. Eur. J. Endocrinol. 2007, 156 (Suppl. 1), S13–S21. [Google Scholar] [CrossRef]
- Dohlman, H.G.; Caron, M.G.; DeBlasi, A.; Frielle, T.; Lefkowitz, R.J. Role of extracellular disulfide-bonded cysteines in the ligand binding function of the beta 2-adrenergic receptor. Biochemistry 1990, 29, 2335–2342. [Google Scholar] [CrossRef]
- Gazi, L.; Nickolls, S.A.; Strange, P.G. Functional coupling of the human dopamine D2 receptor with G alpha i1, G alpha i2, G alpha i3 and G alpha o G proteins: Evidence for agonist regulation of G protein selectivity. Br. J. Pharmacol. 2003, 138, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Spicher, K.; Boulay, G.; Wang, Y.; Birnbaumer, L. Most central nervous system D2 dopamine receptors are coupled to their effectors by Go. Proc. Natl. Acad. Sci. USA 2001, 98, 3577–3582. [Google Scholar] [CrossRef] [Green Version]
- Senogles, S.E.; Spiegel, A.M.; Padrell, E.; Iyengar, R.; Caron, M.G. Specificity of receptor-G protein interactions. Discrimination of Gi subtypes by the D2 dopamine receptor in a reconstituted system. J. Biol. Chem. 1990, 265, 4507–4514. [Google Scholar]
- Montmayeur, J.P.; Guiramand, J.; Borrelli, E. Preferential coupling between dopamine D2 receptors and G-proteins. Mol. Endocrinol. 1993, 7, 161–170. [Google Scholar]
- Zuk, J.; Bartuzi, D.; Matosiuk, D.; Kaczor, A.A. Preferential Coupling of Dopamine D2S and D2L Receptor Isoforms with Gi1 and Gi2 Proteins-In Silico Study. Int. J. Mol. Sci. 2020, 21, 436. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common activation mechanism of class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Chen, K.M.; Clark, M.J.; Hijazi, M.; Kumari, P.; Bai, X.C.; Sunahara, R.K.; Barth, P.; Rosenbaum, D.M. Structure of a D2 dopamine receptor-G-protein complex in a lipid membrane. Nature 2020, 584, 125–129. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Fukunaga, K. Differential subcellular localization of two dopamine D2 receptor isoforms in transfected NG108-15 cells. J. Neurochem. 2003, 85, 1064–1074. [Google Scholar] [CrossRef]
- Kaczor, A.A.; Jorg, M.; Capuano, B. The dopamine D2 receptor dimer and its interaction with homobivalent antagonists: Homology modeling, docking and molecular dynamics. J. Mol. Model 2016, 22, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madras, B.K. History of the discovery of the antipsychotic dopamine D2 receptor: A basis for the dopamine hypothesis of schizophrenia. J. Hist. Neurosci. 2013, 22, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Giros, B.; Sokoloff, P.; Martres, M.P.; Riou, J.F.; Emorine, L.J.; Schwartz, J.C. Alternative splicing directs the expression of two D2 dopamine receptor isoforms. Nature 1989, 342, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Monsma, F.J., Jr.; McVittie, L.D.; Gerfen, C.R.; Mahan, L.C.; Sibley, D.R. Multiple D2 dopamine receptors produced by alternative RNA splicing. Nature 1989, 342, 926–929. [Google Scholar] [CrossRef]
- Dal Toso, R.; Sommer, B.; Ewert, M.; Herb, A.; Pritchett, D.B.; Bach, A.; Shivers, B.D.; Seeburg, P.H. The dopamine D2 receptor: Two molecular forms generated by alternative splicing. EMBO J. 1989, 8, 4025–4034. [Google Scholar] [CrossRef]
- Pelham, H.R. Sorting and retrieval between the endoplasmic reticulum and Golgi apparatus. Curr. Opin. Cell Biol. 1995, 7, 530–535. [Google Scholar] [CrossRef]
- Montmayeur, J.P.; Bausero, P.; Amlaiky, N.; Maroteaux, L.; Hen, R.; Borrelli, E. Differential expression of the mouse D2 dopamine receptor isoforms. FEBS Lett. 1991, 278, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Sedaghat, K.; Nantel, M.F.; Ginsberg, S.; Lalonde, V.; Tiberi, M. Molecular characterization of dopamine D2 receptor isoforms tagged with green fluorescent protein. Mol. Biotechnol. 2006, 34, 1–14. [Google Scholar] [CrossRef]
- Prou, D.; Gu, W.J.; Le Crom, S.; Vincent, J.D.; Salamero, J.; Vernier, P. Intracellular retention of the two isoforms of the D(2) dopamine receptor promotes endoplasmic reticulum disruption. J. Cell Sci. 2001, 114 Pt 19, 3517–3527. [Google Scholar]
- Usiello, A.; Baik, J.H.; Rouge-Pont, F.; Picetti, R.; Dierich, A.; LeMeur, M.; Piazza, P.V.; Borrelli, E. Distinct functions of the two isoforms of dopamine D2 receptors. Nature 2000, 408, 199–203. [Google Scholar] [CrossRef]
- Radl, D.; Chiacchiaretta, M.; Lewis, R.G.; Brami-Cherrier, K.; Arcuri, L.; Borrelli, E. Differential regulation of striatal motor behavior and related cellular responses by dopamine D2L and D2S isoforms. Proc. Natl. Acad. Sci. USA 2018, 115, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, B.G.; Condon, A.F.; Radl, D.; Borrelli, E.; Williams, J.T.; Neve, K.A. Cocaine-induced adaptation of dopamine D2S, but not D2L autoreceptors. eLife 2017, 6, e31924. [Google Scholar] [CrossRef]
- Margeta-Mitrovic, M.; Jan, Y.N.; Jan, L.Y. A trafficking checkpoint controls GABA(B) receptor heterodimerization. Neuron 2000, 27, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Snyder, G.L.; Vanover, K.E. Dopamine Targeting Drugs for the Treatment of Schizophrenia: Past, Present and Future. Curr. Top. Med. Chem. 2016, 16, 3385–3403. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Che, T.; Levit, A.; Shoichet, B.K.; Wacker, D.; Roth, B.L. Structure of the D2 dopamine receptor bound to the atypical antipsychotic drug risperidone. Nature 2018, 555, 269–273. [Google Scholar] [CrossRef]
- Martire, G.; Mottola, G.; Pascale, M.C.; Malagolini, N.; Turrini, I.; Serafini-Cessi, F.; Jackson, M.R.; Bonatti, S. Different fate of a single reporter protein containing KDEL or KKXX targeting signals stably expressed in mammalian cells. J. Biol. Chem. 1996, 271, 3541–3547. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Navarro, N.; Miller, E. Protein sorting at the ER-Golgi interface. J. Cell Biol. 2016, 215, 769–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.U.; Mrzljak, L.; Gutierrez, A.; de la Calle, A.; Goldman-Rakic, P.S. Prominence of the dopamine D2 short isoform in dopaminergic pathways. Proc. Natl. Acad. Sci. USA 1998, 95, 7731–7736. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Aranda, M.F.; Acevedo, M.J.; Gutierrez, A.; Koulen, P.; Khan, Z.U. Role of a Galphai2 protein splice variant in the formation of an intracellular dopamine D2 receptor pool. J. Cell Sci. 2007, 120 Pt 13, 2171–2178. [Google Scholar] [CrossRef] [Green Version]
- Macey, T.A.; Gurevich, V.V.; Neve, K.A. Preferential Interaction between the dopamine D2 receptor and Arrestin2 in neostriatal neurons. Mol. Pharmacol. 2004, 66, 1635–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, G.Y.; Varghese, G.; Chung, H.T.; Trogadis, J.; Seeman, P.; O’Dowd, B.F.; George, S.R. Resistance of the dopamine D2L receptor to desensitization accompanies the up-regulation of receptors on to the surface of Sf9 cells. Endocrinology 1997, 138, 4199–4206. [Google Scholar] [CrossRef] [PubMed]
- Van Craenenbroeck, K.; Clark, S.D.; Cox, M.J.; Oak, J.N.; Liu, F.; Van Tol, H.H. Folding efficiency is rate-limiting in dopamine D4 receptor biogenesis. J. Biol. Chem. 2005, 280, 19350–19357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achour, L.; Labbe-Jullie, C.; Scott, M.G.; Marullo, S. An escort for GPCRs: Implications for regulation of receptor density at the cell surface. Trends Pharmacol. Sci. 2008, 29, 528–535. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.I.; Min, C.; Jung, K.S.; Cheong, S.Y.; Zheng, M.; Cheong, S.J.; Oak, M.H.; Cheong, J.H.; Lee, B.K.; Kim, K.M. The N-terminal region of the dopamine D2 receptor, a rhodopsin-like GPCR, regulates correct integration into the plasma membrane and endocytic routes. Br. J. Pharmacol. 2012, 166, 659–675. [Google Scholar] [CrossRef] [Green Version]
- Fishburn, C.S.; Elazar, Z.; Fuchs, S. Differential glycosylation and intracellular trafficking for the long and short isoforms of the D2 dopamine receptor. J. Biol. Chem. 1995, 270, 29819–29824. [Google Scholar]
- Grunewald, S.; Reilander, H.; Michel, H. In vivo reconstitution of dopamine D2S receptor-mediated G protein activation in baculovirus-infected insect cells: Preferred coupling to Gi1 versus Gi2. Biochemistry 1996, 35, 15162–15173. [Google Scholar] [CrossRef]
- Senogles, S.E. The D2 dopamine receptor isoforms signal through distinct Gi alpha proteins to inhibit adenylyl cyclase. A study with site-directed mutant Gi alpha proteins. J. Biol. Chem. 1994, 269, 23120–23127. [Google Scholar]
- Choi, D.S.; Wang, D.; Tolbert, L.; Sadee, W. Basal signaling activity of human dopamine D2L receptor demonstrated with an ecdysone-inducible mammalian expression system. J. Neurosci. Methods 2000, 94, 217–225. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, R.; Sasaoka, T.; Tonegawa, S.; Kung, M.P.; Sankoorikal, E.B. Dopamine D2 long receptor-deficient mice display alterations in striatum-dependent functions. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 8305–8314. [Google Scholar] [CrossRef] [Green Version]
- Welter, M.; Vallone, D.; Samad, T.A.; Meziane, H.; Usiello, A.; Borrelli, E. Absence of dopamine D2 receptors unmasks an inhibitory control over the brain circuitries activated by cocaine. Proc. Natl. Acad. Sci. USA 2007, 104, 6840–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Hranilovic, D.; Fetsko, L.A.; Bucan, M.; Wang, Y. Dopamine D2S and D2L receptors may differentially contribute to the actions of antipsychotic and psychotic agents in mice. Mol. Psychiatry 2002, 7, 1075–1082. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Filipeanu, C.M.; Duvernay, M.T.; Wu, G. Regulation of G protein-coupled receptor export trafficking. Biochim. Biophys. Acta 2007, 1768, 853–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doly, S.; Shirvani, H.; Gata, G.; Meye, F.J.; Emerit, M.B.; Enslen, H.; Achour, L.; Pardo-Lopez, L.; Yang, S.K.; Armand, V.; et al. GABAB receptor cell-surface export is controlled by an endoplasmic reticulum gatekeeper. Mol. Psychiatry 2016, 21, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wu, G. Mechanisms of the anterograde trafficking of GPCRs: Regulation of AT1R transport by interacting proteins and motifs. Traffic 2019, 20, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Milligan, G. The role of dimerisation in the cellular trafficking of G-protein-coupled receptors. Curr. Opin. Pharmacol. 2010, 10, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Regulation of post-Golgi traffic of G protein-coupled receptors. Sub-Cell. Biochem. 2012, 63, 83–95. [Google Scholar]
- Spang, A. Retrograde traffic from the Golgi to the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2013, 5, a013391. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Nilsson, T.; Peterson, P.A. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990, 9, 3153–3162. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Pelham, H.R. A C-terminal signal prevents secretion of luminal ER proteins. Cell 1987, 48, 899–907. [Google Scholar] [CrossRef]
- Jin, H.; Komita, M.; Aoe, T. The Role of BiP Retrieval by the KDEL Receptor in the Early Secretory Pathway and its Effect on Protein Quality Control and Neurodegeneration. Front. Mol. Neurosci. 2017, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Rothman, J.E.; Wieland, F.T. Protein sorting by transport vesicles. Science 1996, 272, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Cukierman, E.; Huber, I.; Rotman, M.; Cassel, D. The ARF1 GTPase-activating protein: Zinc finger motif and Golgi complex localization. Science 1995, 270, 1999–2002. [Google Scholar] [CrossRef]
- Aoe, T.; Huber, I.; Vasudevan, C.; Watkins, S.C.; Romero, G.; Cassel, D.; Hsu, V.W. The KDEL receptor regulates a GTPase-activating protein for ADP-ribosylation factor 1 by interacting with its non-catalytic domain. J. Biol. Chem. 1999, 274, 20545–20549. [Google Scholar] [CrossRef] [Green Version]
- Boehmler, W.; Obrecht-Pflumio, S.; Canfield, V.; Thisse, C.; Thisse, B.; Levenson, R. Evolution and expression of D2 and D3 dopamine receptor genes in zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 230, 481–493. [Google Scholar] [CrossRef]
- Kokubun, H.; Jin, H.; Aoe, T. Pathogenic Effects of Impaired Retrieval between the Endoplasmic Reticulum and Golgi Complex. Int. J. Mol. Sci. 2019, 20, 5614. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, M.; Muniz, M.; Hidalgo, J.; Vega, L.; Martin, M.E.; Velasco, A. The retrieval function of the KDEL receptor requires PKA phosphorylation of its C-terminus. Mol. Biol. Cell 2003, 14, 4114–4125. [Google Scholar] [CrossRef]
- Jackson, L.P.; Lewis, M.; Kent, H.M.; Edeling, M.A.; Evans, P.R.; Duden, R.; Owen, D.J. Molecular basis for recognition of dilysine trafficking motifs by COPI. Dev. Cell 2012, 23, 1255–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letourneur, F.; Gaynor, E.C.; Hennecke, S.; Demolliere, C.; Duden, R.; Emr, S.D.; Riezman, H.; Cosson, P. Coatomer is essential for retrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell 1994, 79, 1199–1207. [Google Scholar] [CrossRef]
- Vrecl, M.; Anderson, L.; Hanyaloglu, A.; McGregor, A.M.; Groarke, A.D.; Milligan, G.; Taylor, P.L.; Eidne, K.A. Agonist-induced endocytosis and recycling of the gonadotropin-releasing hormone receptor: Effect of beta-arrestin on internalization kinetics. Mol. Endocrinol. 1998, 12, 1818–1829. [Google Scholar] [PubMed] [Green Version]
- Pagano, A.; Rovelli, G.; Mosbacher, J.; Lohmann, T.; Duthey, B.; Stauffer, D.; Ristig, D.; Schuler, V.; Meigel, I.; Lampert, C.; et al. C-terminal interaction is essential for surface trafficking but not for heteromeric assembly of GABA(b) receptors. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 1189–1202. [Google Scholar] [CrossRef] [Green Version]
- Kubale, V.; Pogacnik, A.; Schwartz, W.T.; Vrecl, M. Seven transmembrane receptors (7TM) in the view of dimerization and experimental methods to study their dimerization and cross-talk. Slov. Vet. Res. 2008, 45, 89–102. [Google Scholar]
- Kuner, R.; Kohr, G.; Grunewald, S.; Eisenhardt, G.; Bach, A.; Kornau, H.C. Role of heteromer formation in GABAB receptor function. Science 1999, 283, 74–77. [Google Scholar] [CrossRef]
- Filipeanu, C.M.; Pullikuth, A.K.; Guidry, J.J. Molecular determinants of the human alpha2C-adrenergic receptor temperature-sensitive intracellular traffic. Mol. Pharmacol. 2015, 87, 792–802. [Google Scholar] [CrossRef] [Green Version]
- Nasu-Nishimura, Y.; Hurtado, D.; Braud, S.; Tang, T.T.; Isaac, J.T.; Roche, K.W. Identification of an endoplasmic reticulum-retention motif in an intracellular loop of the kainate receptor subunit KA2. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 7014–7021. [Google Scholar] [CrossRef] [Green Version]
- Hermosilla, R.; Schulein, R. Sorting functions of the individual cytoplasmic domains of the G protein-coupled vasopressin V(2) receptor in Madin Darby canine kidney epithelial cells. Mol. Pharmacol. 2001, 60, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Kubale, V.; Blagotinsek, K.; Nohr, J.; Eidne, K.A.; Vrecl, M. The Conserved Arginine Cluster in the Insert of the Third Cytoplasmic Loop of the Long Form of the D(2) Dopamine Receptor (D2L-R) Acts as an Intracellular Retention Signal. Int. J. Mol. Sci. 2016, 17, 1152. [Google Scholar] [CrossRef] [Green Version]
- Shioda, N.; Takeuchi, Y.; Fukunaga, K. Advanced research on dopamine signaling to develop drugs for the treatment of mental disorders: Proteins interacting with the third cytoplasmic loop of dopamine D2 and D3 receptors. J. Pharmacol. Sci. 2010, 114, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Navarro, G.; Ferre, S.; Cordomi, A.; Moreno, E.; Mallol, J.; Casado, V.; Cortes, A.; Hoffmann, H.; Ortiz, J.; Canela, E.I.; et al. Interactions between intracellular domains as key determinants of the quaternary structure and function of receptor heteromers. J. Biol. Chem. 2010, 285, 27346–27359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukasiewicz, S.; Faron-Gorecka, A.; Dobrucki, J.; Polit, A.; Dziedzicka-Wasylewska, M. Studies on the role of the receptor protein motifs possibly involved in electrostatic interactions on the dopamine D1 and D2 receptor oligomerization. FEBS J. 2009, 276, 760–775. [Google Scholar] [CrossRef] [PubMed]
- Lukasiewicz, S.; Polit, A.; Kedracka-Krok, S.; Wedzony, K.; Mackowiak, M.; Dziedzicka-Wasylewska, M. Hetero-dimerization of serotonin 5-HT(2A) and dopamine D(2) receptors. Biochim. Biophys. Acta 2010, 1803, 1347–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Likhite, N.; Jackson, C.A.; Liang, M.S.; Krzyzanowski, M.C.; Lei, P.; Wood, J.F.; Birkaya, B.; Michaels, K.L.; Andreadis, S.T.; Clark, S.D.; et al. The protein arginine methyltransferase PRMT5 promotes D2-like dopamine receptor signaling. Sci. Signal. 2015, 8, ra115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Gouw, M.; Michael, S.; Diakogianni, A.; Valverde, J.A.; Bukirova, D.; Calyseva, J.; Palopoli, N.; Davey, N.E.; Chemes, L.B.; et al. ELM-the eukaryotic linear motif resource in 2020. Nucleic Acids Res. 2020, 48, D296–D306. [Google Scholar] [CrossRef] [Green Version]
- Duvernay, M.T.; Filipeanu, C.M.; Wu, G. The regulatory mechanisms of export trafficking of G protein-coupled receptors. Cell. Signal. 2005, 17, 1457–1465. [Google Scholar] [CrossRef]
- Bermak, J.C.; Li, M.; Bullock, C.; Zhou, Q.Y. Regulation of transport of the dopamine D1 receptor by a new membrane-associated ER protein. Nat. Cell Biol. 2001, 3, 492–498. [Google Scholar] [CrossRef]
- Kabbani, N.; Woll, M.P.; Nordman, J.C.; Levenson, R. Dopamine receptor interacting proteins: Targeting neuronal calcium sensor-1/D2 dopamine receptor interaction for antipsychotic drug development. Curr. Drug Targets 2012, 13, 72–79. [Google Scholar] [CrossRef]
- Sencanski, M.; Glisic, S.; Snajder, M.; Veljkovic, N.; Poklar Ulrih, N.; Mavri, J.; Vrecl, M. Computational design and characterization of nanobody-derived peptides that stabilize the active conformation of the beta2-adrenergic receptor (beta2-AR). Sci. Rep. 2019, 9, 16555. [Google Scholar] [CrossRef] [Green Version]
- Mandic, M.; Drinovec, L.; Glisic, S.; Veljkovic, N.; Nohr, J.; Vrecl, M. Demonstration of a direct interaction between beta2-adrenergic receptor and insulin receptor by BRET and bioinformatics. PLoS ONE 2014, 9, e112664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.Y.; Chu, X.P.; Mao, L.M.; Wang, M.; Lan, H.X.; Li, M.H.; Zhang, G.C.; Parelkar, N.K.; Fibuch, E.E.; Haines, M.; et al. Modulation of D2R-NR2B interactions in response to cocaine. Neuron 2006, 52, 897–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Fu, W.; Xie, J.; Luo, H.; Sells, S.F.; Geddes, J.W.; Bondada, V.; Rangnekar, V.M.; Mattson, M.P. Par-4 is a mediator of neuronal degeneration associated with the pathogenesis of Alzheimer disease. Nat. Med. 1998, 4, 957–962. [Google Scholar] [CrossRef]
- Park, S.K.; Nguyen, M.D.; Fischer, A.; Luke, M.P.; Affar, E.B.; Dieffenbach, P.B.; Tseng, H.C.; Shi, Y.; Tsai, L.H. Par-4 links dopamine signaling and depression. Cell 2005, 122, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binda, A.V.; Kabbani, N.; Lin, R.; Levenson, R. D2 and D3 dopamine receptor cell surface localization mediated by interaction with protein 4.1N. Mol. Pharmacol. 2002, 62, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Karpa, K.; Kabbani, N.; Goldman-Rakic, P.; Levenson, R. Dopamine D2 and D3 receptors are linked to the actin cytoskeleton via interaction with filamin A. Proc. Natl. Acad. Sci. USA 2001, 98, 5258–5263. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.J.; Pei, L.; Moszczynska, A.; Vukusic, B.; Fletcher, P.J.; Liu, F. Dopamine transporter cell surface localization facilitated by a direct interaction with the dopamine D2 receptor. EMBO J. 2007, 26, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Navarro, G.; Aymerich, M.S.; Marcellino, D.; Cortes, A.; Casado, V.; Mallol, J.; Canela, E.I.; Agnati, L.; Woods, A.S.; Fuxe, K.; et al. Interactions between calmodulin, adenosine A2A, and dopamine D2 receptors. J. Biol. Chem. 2009, 284, 28058–28068. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.S.; Marcellino, D.; Jackson, S.N.; Franco, R.; Ferre, S.; Agnati, L.F.; Fuxe, K. How calmodulin interacts with the adenosine A(2A) and the dopamine D(2) receptors. J. Proteome Res. 2008, 7, 3428–3434. [Google Scholar] [CrossRef] [Green Version]
- Hanson, P.I.; Otto, H.; Barton, N.; Jahn, R. The N-ethylmaleimide-sensitive fusion protein and alpha-SNAP induce a conformational change in syntaxin. J. Biol. Chem. 1995, 270, 16955–16961. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Li, L.; Pei, L.; Vukusic, B.; Van Tol, H.H.; Lee, F.J.; Wan, Q.; Liu, F. Protein-protein coupling/uncoupling enables dopamine D2 receptor regulation of AMPA receptor-mediated excitotoxicity. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 4385–4395. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.D.; Oxford, G.S.; Milgram, S.L. Association of the D2 dopamine receptor third cytoplasmic loop with spinophilin, a protein phosphatase-1-interacting protein. J. Biol. Chem. 1999, 274, 19894–19900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.Z.; Imachi, H.; Lyu, J.Y.; Fukunaga, K.; Sato, S.; Ibata, T.; Kobayashi, T.; Yoshimoto, T.; Kikuchi, F.; Dong, T.; et al. Prolactin regulatory element-binding protein is involved in suppression of the adiponectin gene in vivo. J. Endocrinol. Investig. 2017, 40, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Kim, M.Y.; Kim, T.H.; Min, D.K.; Yang, G.E.; Ahn, Y.H. Prolactin regulatory element-binding (PREB) protein regulates hepatic glucose homeostasis. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2097–2107. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Murao, K.; Imachi, H.; Li, J.; Nishiuchi, T.; Dobashi, H.; Hosomi, N.; Masugata, H.; Zhang, G.X.; Iwama, H.; et al. The transcription factor prolactin regulatory element-binding protein mediates prolactin transcription induced by thyrotropin-releasing hormone in GH3 cells. Endocrine 2010, 38, 53–59. [Google Scholar] [CrossRef]
- Zhang, W.; Murao, K.; Imachi, H.; Iwama, H.; Chen, K.; Fei, Z.; Zhang, X.; Ishida, T.; Tamiya, T. Suppression of prolactin expression by cabergoline requires prolactin regulatory element-binding protein (PREB) in GH3 cells. Horm. Metab. Res. 2010, 42, 557–561. [Google Scholar] [CrossRef]
- Wimalasena, D.S.; Wimalasena, K. Kinetic evidence for channeling of dopamine between monoamine transporter and membranous dopamine-beta-monooxygenase in chromaffin granule ghosts. J. Biol. Chem. 2004, 279, 15298–15304. [Google Scholar] [CrossRef] [Green Version]
- Mochida, G.H.; Mahajnah, M.; Hill, A.D.; Basel-Vanagaite, L.; Gleason, D.; Hill, R.S.; Bodell, A.; Crosier, M.; Straussberg, R.; Walsh, C.A. A truncating mutation of TRAPPC9 is associated with autosomal-recessive intellectual disability and postnatal microcephaly. Am. J. Hum. Genet. 2009, 85, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Shioda, N. Dopamine D2L receptor-interacting proteins regulate dopaminergic signaling. J. Pharmacol. Sci. 2017, 135, 51–54. [Google Scholar] [CrossRef]
- Shioda, N.; Yamamoto, Y.; Watanabe, M.; Binas, B.; Owada, Y.; Fukunaga, K. Heart-type fatty acid binding protein regulates dopamine D2 receptor function in mouse brain. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 3146–3155. [Google Scholar] [CrossRef]
- Yabuki, Y.; Takahata, I.; Matsuo, K.; Owada, Y.; Fukunaga, K. Ramelteon Improves Post-traumatic Stress Disorder-Like Behaviors Exhibited by Fatty Acid-Binding Protein 3 Null Mice. Mol. Neurobiol. 2018, 55, 3577–3591. [Google Scholar] [CrossRef]
- Brismar, H.; Asghar, M.; Carey, R.M.; Greengard, P.; Aperia, A. Dopamine-induced recruitment of dopamine D1 receptors to the plasma membrane. Proc. Natl. Acad. Sci. USA 1998, 95, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Holtback, U.; Brismar, H.; DiBona, G.F.; Fu, M.; Greengard, P.; Aperia, A. Receptor recruitment: A mechanism for interactions between G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 1999, 96, 7271–7275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, A.M.; Liu, Y.; Vidensky, S.; Maier, S.; Jung, E.; Farhan, H.; Robinson, M.B.; Sitte, H.H.; Rothstein, J.D. The endoplasmic reticulum exit of glutamate transporter is regulated by the inducible mammalian Yip6b/GTRAP3-18 protein. J. Biol. Chem. 2008, 283, 6175–6183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupre, D.J.; Robitaille, M.; Richer, M.; Ethier, N.; Mamarbachi, A.M.; Hebert, T.E. Dopamine receptor-interacting protein 78 acts as a molecular chaperone for Ggamma subunits before assembly with Gbeta. J. Biol. Chem. 2007, 282, 13703–13715. [Google Scholar] [CrossRef] [Green Version]
- Bowers, M.S.; McFarland, K.; Lake, R.W.; Peterson, Y.K.; Lapish, C.C.; Gregory, M.L.; Lanier, S.M.; Kalivas, P.W. Activator of G protein signaling 3: A gatekeeper of cocaine sensitization and drug seeking. Neuron 2004, 42, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Rassu, M.; Del Giudice, M.G.; Sanna, S.; Taymans, J.M.; Morari, M.; Brugnoli, A.; Frassineti, M.; Masala, A.; Esposito, S.; Galioto, M.; et al. Role of LRRK2 in the regulation of dopamine receptor trafficking. PLoS ONE 2017, 12, e0179082. [Google Scholar] [CrossRef] [PubMed]
- Itokawa, M.; Arinami, T.; Toru, M. Advanced research on dopamine signaling to develop drugs for the treatment of mental disorders: Ser311Cys polymorphisms of the dopamine D2-receptor gene and schizophrenia. J. Pharmacol. Sci. 2010, 114, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Wainwright, M.S.; Comeron, J.M.; Saitou, N.; Sanders, A.R.; Gelernter, J.; Gejman, P.V. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA stability and synthesis of the receptor. Hum. Mol. Genet. 2003, 12, 205–216. [Google Scholar] [CrossRef]
- Guiramand, J.; Montmayeur, J.P.; Ceraline, J.; Bhatia, M.; Borrelli, E. Alternative splicing of the dopamine D2 receptor directs specificity of coupling to G-proteins. J. Biol. Chem. 1995, 270, 7354–7358. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, R.; Hofer, A.; Grapengiesser, A.; Gasser, T.; Kupsch, A.; Roots, I.; Brockmoller, J. L -dopa-induced adverse effects in PD and dopamine transporter gene polymorphism. Neurology 2003, 60, 1750–1755. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D.; Urbanek, M.; Guenther, D.; Robin, R.; Long, J.C. Linkage and association of a functional DRD2 variant [Ser311Cys] and DRD2 markers to alcoholism, substance abuse and schizophrenia in Southwestern American Indians. Am. J. Med. Genet. 1997, 74, 386–394. [Google Scholar] [CrossRef]
- Senogles, S.E.; Heimert, T.L.; Odife, E.R.; Quasney, M.W. A region of the third intracellular loop of the short form of the D2 dopamine receptor dictates Gi coupling specificity. J. Biol. Chem. 2004, 279, 1601–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| ELM Name | Instances (Matched Sequence) | AA Position | ELM Description | Cell Compartment | Pattern | Probability (×103) |
|---|---|---|---|---|---|---|
| TRG_ER_diArg_1 | RRRR RRR RRKR RKRV RRR RRV | 217–220 218–220 219–222 220–223 267–269 268–270 | The di-Arg ER retention motif is defined by two consecutive RR residues or with a single residue insertion (RXR). The motif is completed by an adjacent hydrophobic/arginine residue, which may be on either side of the R pair. | ER membrane, integral protein, ER-GA transport vesicle membrane, ER membrane, GA-ER transport vesicle membrane, rough ER, ER | ([LIVMFYWPR]R[^YFWDE]{0,1}R)|(R[^YFWDE]{0,1}R[LIVMFYWPR]) | 5.37 |
| CLV_NRD_NRD_1 | RRK RRV RRA RRK FRK | 219–221 268–270 274–276 360–362 433–435 | N-Arg dibasic convertase (NRD/Nardilysin) cleavage site (X-|-R-K or R-|-R-X). | extracellular, GA, cell surface | (.RK)|(RR[^KR]) | 7.47 |
| CLV_PCSK_FUR_1 | RRRK RRKRV | 217–221 219–223 | Furin (PACE) cleavage site (R-X-[RK]-R-|-X). | extracellular, GA, GA membrane | R.[RK]R. | 50.09 |
| CLV_PCSK_KEX2_1 | KRR RRR RRR RRK KRV KRS RRR RRV RRA RRK | 149–151 217–219 218–220 219–221 221–223 226–228 267–269 268–270 274–276 360–362 | Yeast kexin 2 cleavage site (K-R-|-X or R-R-|-X). | extracellular, GA | [KR]R. | 7.97 |
| CLV_PCSK_PC1ET2_1 | KRR KRV KRS | 149–151 221–223 226–228 | NEC1/NEC2 cleavage site (K-R-|-X). | extracellular, GA, GA membrane | KR. | 3.90 |
| CLV_PCSK_PC7_1 | RYSSKRR RRRRKRV RVNTKRS | 145–151 217–223 222–228 | Proprotein convertase 7 (PC7, PCSK7) cleavage site (R-X-X-X-[RK]-R-|-X). | extracellular, GA, GA membrane | R...[KR]R. | 50.09 |
| CLV_PCSK_SKI1_1 | KIAKI KKATQ RKAFL KAFLK | 336–340 369–373 434–438 435–439 | Subtilisin/kexin isozyme-1 (SKI1) cleavage site ([RK]-X-[hydrophobic]-[LTKF]-|-X). | ER lumen, ER GA, extracellular | [RK].[AILMFV][LTKF]. | 6.82 |
| LIG_deltaCOP1_diTrp_1 | EWKF | 99–105 | Tryptophan-based motifs enable targeting of the tethering and (dis)assembly factors to the C-terminal mu homology domain (MHD) of the coatomer subunit delta, delta-COP. | ER membrane GA membrane, COPI coated vesicle membrane, cytosol, COPI vesicle coat, transport vesicle | [DE]{1,3}.{0,2}W.{1,6}[WF] | 50.10 |
| LIG_LIR_Gen_1 | EWKFSRI | 99–105 | Canonical LIR motif that binds to Atg8 protein family members to mediate processes involved in autophagy. | cytosol, cytoplasmic side of late endosome membrane | [EDST].{0,2}[WFY][^RKPG][^PG][ILV] | 3.06 |
| LIG_LIR_Nem_3 | EWKFSRI TRYSPI | 99–105 293–298 | Nematode-specific variant of the canonical LIR motif that binds to Atg8 protein family members to mediate processes involved in autophagy. | cytosol, cytoplasmic side of late endosome membrane | [EDST].{0,2}[WFY]..[ILVFY] | 6.36 |
| MOD_N-GLC_1 | EWKFSRI TRYSPI | 99–105 293–298 | Generic motif for N-glycosylation. It was shown that Trp, Asp, and Glu are uncommon before the Ser/Thr position. Efficient glycosylation usually occurs when ~60 residues or more separate the glycosylation acceptor site from the C-terminus. | extracellular, GA, ER | [EDST].{0,2}[WFY]..[ILVFY] | 5.02 |
| MOD_N-GLC_2 | NEC | 180–182 | Atypical motif for N-glycosylation site. Examples are Human CD69, which is uniquely glycosylated at typical (Asn-X-Ser/Thr) and atypical (Asn-X-Cys) motifs, beta protein C. | extracellular, GA, ER | (N)[^P]C | 29.7 |
| TRG_ENDOCYTIC_2 | YTAV YSPI | 133–136 295–298 | Tyrosine-based sorting signal responsible for the interaction with mu subunit of AP (Adaptor Protein) complex. | PM, clathrin-coated endocytic vesicle, cytosol | Y.[LMVIF] | 2.59 |
| Receptor | AA Residue | Location | Mutation | Reference |
|---|---|---|---|---|
| drd2_human | 249 | ICL3 | D => V | Guiramand, J. et al. (1995) [110] |
| drd2_human | 264 | ICL3 | P => G | Guiramand, J. et al. (1995) [110] |
| drd2_human | 310 | ICL3 | P => S | Kaiser, R. et al. (2003) [111] |
| drd2_human | 311 | ICL3 | S => C | Kaiser, R. et al. (2003) [111] |
| drd2_human | 311 | ICL3 | S => C | Goldman D et al. (1997) [112] |
| drd2_mouse | 251 | ICL3 | K => V | Guiramand, J. et al. (1995) [110] |
| drd2_mouse | 271 | ICL3 | D => V | Guiramand, J. et al. (1995) [110] |
| drd2_rat | 233 | ICL3 | R => G | Senogles, SE. et al. (2004) [113] |
| drd2_rat | 234 | ICL3 | A => T | Senogles, SE. et al. (2004) [113] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blagotinšek Cokan, K.; Mavri, M.; Rutland, C.S.; Glišić, S.; Senćanski, M.; Vrecl, M.; Kubale, V. Critical Impact of Different Conserved Endoplasmic Retention Motifs and Dopamine Receptor Interacting Proteins (DRIPs) on Intracellular Localization and Trafficking of the D2 Dopamine Receptor (D2-R) Isoforms. Biomolecules 2020, 10, 1355. https://doi.org/10.3390/biom10101355
Blagotinšek Cokan K, Mavri M, Rutland CS, Glišić S, Senćanski M, Vrecl M, Kubale V. Critical Impact of Different Conserved Endoplasmic Retention Motifs and Dopamine Receptor Interacting Proteins (DRIPs) on Intracellular Localization and Trafficking of the D2 Dopamine Receptor (D2-R) Isoforms. Biomolecules. 2020; 10(10):1355. https://doi.org/10.3390/biom10101355
Chicago/Turabian StyleBlagotinšek Cokan, Kaja, Maša Mavri, Catrin Sian Rutland, Sanja Glišić, Milan Senćanski, Milka Vrecl, and Valentina Kubale. 2020. "Critical Impact of Different Conserved Endoplasmic Retention Motifs and Dopamine Receptor Interacting Proteins (DRIPs) on Intracellular Localization and Trafficking of the D2 Dopamine Receptor (D2-R) Isoforms" Biomolecules 10, no. 10: 1355. https://doi.org/10.3390/biom10101355