
Krill Oil and Its Bioactive Components as a Potential Therapy for Inflammatory Bowel Disease: Insights from In Vivo and In Vitro Studies

Abstract
:1. Introduction
2. Properties of Krill Oil
3. The Intestinal Barrier
3.1. The Intestinal Barrier and IBD
3.2. Protective Roles of Krill Oil in Intestinal Barrier Impairment Associated with Intestinal Inflammation
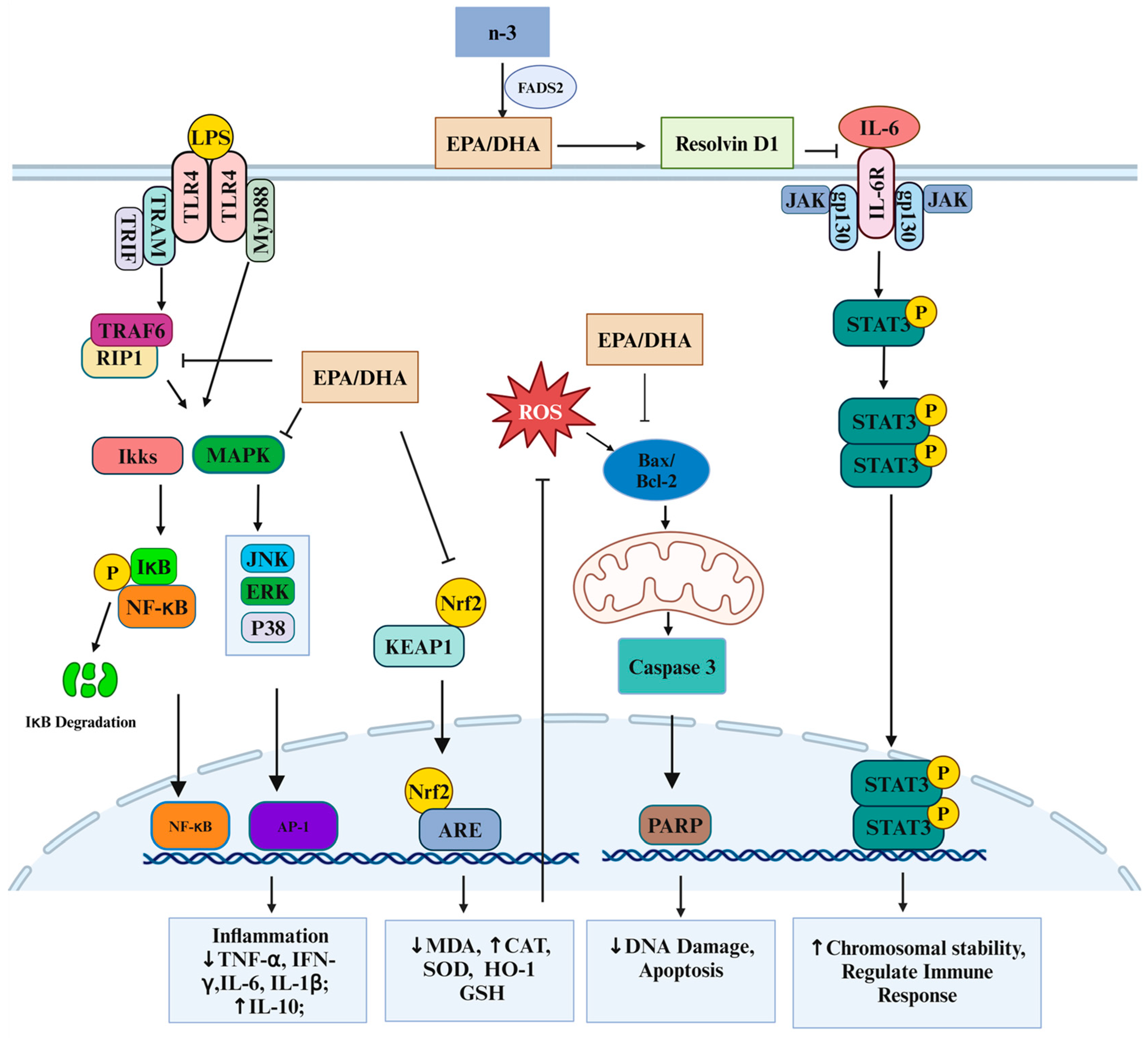
4. Immune Response in Intestinal Inflammation
Anti-Inflammatory Effects of Krill Oil in Intestinal Inflammation
5. Oxidative Stress
5.1. Oxidative Stress and IBD
5.2. Antioxidant Effects of Krill Oil in Inflammatory Conditions
6. Modulation of Gut Microbiota by Krill Oil
7. Clinical Studies Investigating the Effects of Fish Oil and the Status of Krill Oil on IBD

{kind=link}
| Animal Models | Inducer | Treatment | Concentration | Treatment Method | Duration | Effects | Reference |
|---|---|---|---|---|---|---|---|
| DSS-induced colitis in 24 male BALB/c mice | 3.5% DSS in drinking water | KO | Low: 0.25 g/kg High: 0.5 g/kg | Gavage | 21 days: 14-day treatment prior to and 7-day co-treatment with DSS | 0.25 g/kg KO: NO change in body weight and colon length, IL-6,TNF-α, LPS, DAO ↓Histological damage, ↑ZO-1, ↑Occludin, ↓p-p65, ↓p-IκB-α 0.5 g/kg KO: ↓Body weight loss, ↓Disease activity index, ↑Colon length, ↓Histological damage, ↓IL-6, ↓TNF-α, ↑ZO-1, ↑Occludin, ↑Claudin-1, ↓LPS, ↓DAO, ↓p-p65, ↓p- IκB-α, ↑Gut microbiota uniformity and richness, ↑Firmicutes to Bacteroidetes ratios, ↑SCFAs | [64] |
| DSS-induced colitis in 30 male Wistar rat | 5% DSS in drinking water | KO | 5% KO | Mixed with diets | 28 days: 21-day treatment prior to and 7-day co-treatment with DSS | ↓Disease activity index, ↑Colon length, ↓Histological damage, ↓TNF-α, ↓KC/GRO, ↓IL-1β, ↑PGE2, ↑PGE3, ↑PGD2, ↑EPA, ↑DHA, ↓n-6 FAs, ↓GSA, ↓CEL, ↓CML, ↑Ppargc1 a, ↑Pparg, ↑Sirt 1, ↑TLR4, ↓Nos2, ↓IL-6, ↓KC/GRO, ↓Ptgs, ↓Ptgs2 | [78] |
| DSS-induced colitis in 20 female C57BL/6 mice | 3% DSS in drinking water | KLD (KO+VitD3+LR Lactobacillus reuteri) | 16 mg/per mouse | Gavage | 7 days: co-treatment with DSS | ↓Body weight loss, ↑Colon length, ↓Histological damage, ↓TNF-α, ↓IL-1β, ↓IL-6, ↑IL-10 | [134] |
| DSS-induced colitis in 48 male C57BL/6 mice | 2% DSS in drinking water | KO liposomes | 300 μM/kg | Gavage | 9 days: 4-day treatment prior to and 5-day co-treatment with DSS | ↓Disease activity index, ↑Colon length, ↓Histological damage, ↓TNF-α, ↓IL-6, ↓LPS | [65] |
| DSS-induced colitis in 50 male C57BL/6 mice | 3% DSS in drinking water | Antarctic krill phospholipids (APL) | 10, 25, 50 mg/kg | Gavage | 7 days: co-treatment with DSS | 10 mg/kg APL: No change, Body weight loss, Colon weight, Histological damage, Disease activity index, TNF-α, IL-6, ZO-1, ↑Occludin 25 and 50 mg/kg APL: ↓Body weight loss, ↑Colon weight, ↓Histological damage, ↓Disease activity index, ↓TNF-α,↓IL-6, ↑ZO-1, ↑Occludin, 50 mg/kg APL: No change in Chao index of OTU level, ↑Firmicutes/Bacteroidetes ratio, ↑SCFAs (Acetic acid; Propionic acid; Butyric acid) | [79] |
| 40 parasitic whipworm T. suis-infected pigs | Parasitic whipworm T. suis eggs by feeding | KO | 1.5 g daily | Mixed in a freshly made sugar-coated cookie dough ball | 28 days: 7-day treatment prior to and 21-day treatment co-treatment with Parasitic whipworm T. suis | ↓Crypt length, ↑Goblet cell numbers, ↑Histological scores, ↑Microbial alpha diversity indices, ↑Firmicutes to Bacteroidetes ratios, ↓Rickettsiales abundance, ↓Lactobacillus vaginalis species abundance, ↓L-histidine decarboxylase and gut histamine, ↓1-methylhistamine and N-acetylhistamine | [63] |
| 30 C. rodentium infected male C3H/HeNCr mice | C. rodentium (bacteria) by oral gavage | KO | 1.5 mg daily | Gavage | 17 days: 5-day treatment prior to and 12-day co-treatment with C. rodentium | ↓Body weight loss, ↓Histological damage, ↓Load of C. rodentium, improved the microbial dysbiosis index, ↓Lactobacillus reuteri, L. vaginalis, Clostridium perfringens abundance, ↓ IL-1β, ↓TNF, ↓IL-12B, ↓IL-17A, ↓IL-22, ↓CCL2 | [63] |
| DSS-induced colitis in 50 C57BL/6 mice | 3% DSS dissolved in physiological saline and free drinking | KO-HIPPE (high internal phase Pickering emulsion) | 200 mg/kg 400 mg/kg 800 mg/kg | Gavage | 9 days co-treatment with DSS | 200 mg/kg KO-HIPPE: ↑Colon length, ↓Histological damage, ↓TNF-α,↓IL-6, MDA, SOD 400 mg/kg KO-HIPPE: ↑Colon length (No significant difference), ↓Histological damage, ↑Occludin, ↓TNF-α,↓IL-6, MDA, ↑GSH, SOD 800 mg/kg KO-HIPPE: ↑Colon length, ↓Histological damage, ↓TNF-α,↓IL-6, MDA, | [135] |
| 36 C. rodentium infected male C3H/HeNCr mice | C. rodentium (bacteria) by oral gavage | KO in water emulsion | 40 mg KO/Body weight (kg) | Gavage | 26 days: 7-day treatment prior to and 19-day co-treatment with C. rodentium | ↓ Colon index (Colon weight/body weight), Crypt length, IL-1β, Pparg, Gzmk, histamine ↑leap2, Gas7, Muc6, abundance of butyrate producing taxa (Roseburia and Clostridium), microbial interactions, oleic acid, ricinoleic acid, 4-(2-amino-3-hydroxyphenyl)-2,4-dioxobutanoic acid, γ-glutamylglutamic acid, 5-hydroxyindoleacetic acid, Anaeroplasmataceae, Clostridiaceae, Christensenellaceae (family level) Enriched pathways: cGMP-PKG signalling, calcium signalling, phospholipase D signalling, gap junction, and purine metabolism, pathways related to secretion | [66] |
8. Beneficial Effects of EPA and DHA in Intestinal Inflammation
9. Beneficial Effects of Astaxanthin in Intestinal Inflammation
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.R.; Rodriguez, J.R. Clinical presentation of Crohn’s, ulcerative colitis, and indeterminate colitis: Symptoms, extraintestinal manifestations, and disease phenotypes. Semin. Pediatr. Surg. 2017, 26, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. Ulcerative colitis. Nat. Rev. Dis. Prim. 2020, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Roda, G.; Chien Ng, S.; Kotze, P.G.; Argollo, M.; Panaccione, R.; Spinelli, A.; Kaser, A.; Peyrin-Biroulet, L.; Danese, S. Crohn’s disease. Nat. Rev. Dis. Prim. 2020, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Villanacci, V.; Reggiani-Bonetti, L.; Salviato, T.; Leoncini, G.; Cadei, M.; Albarello, L.; Caputo, A.; Aquilano, M.C.; Battista, S.; Parente, P. Histopathology of IBD Colitis. A practical approach from the pathologists of the Italian Group for the study of the gastrointestinal tract (GIPAD). Pathologica 2021, 113, 39–53. [Google Scholar] [CrossRef]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, M.A.; Caruso, A. The Gut Microbiome in Psoriasis and Crohn’s Disease: Is Its Perturbation a Common Denominator for Their Pathogenesis? Vaccines 2022, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, C.A.; Gabbiadini, R.; Dal Buono, A.; Quadarella, A.; De Marco, A.; Repici, A.; Bezzio, C.; Simonetta, E.; Aliberti, S.; Armuzzi, A. Lung Involvement in Inflammatory Bowel Diseases: Shared Pathways and Unwanted Connections. J. Clin. Med. 2023, 12, 6419. [Google Scholar] [CrossRef] [PubMed]
- Jucan, A.E.; Gavrilescu, O.; Dranga, M.; Popa, I.V.; Mihai, B.M.; Prelipcean, C.C.; Mihai, C. Ischemic Heart Disease in Patients with Inflammatory Bowel Disease: Risk Factors, Mechanisms and Prevention. Life 2022, 12, 1113. [Google Scholar] [CrossRef]
- Abu-Freha, N.; Cohen, B.; Gordon, M.; Weissmann, S.; Kestenbaum, E.H.; Vosko, S.; Abu-Tailakh, M.; Ben-Shoshan, L.; Cohen, D.L.; Shirin, H. Colorectal cancer among inflammatory bowel disease patients: Risk factors and prevalence compared to the general population. Front. Med. 2023, 10, 1225616. [Google Scholar] [CrossRef]
- Santana, P.T.; Rosas, S.L.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S.P. Dysbiosis in Inflammatory Bowel Disease: Pathogenic Role and Potential Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 3464. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, K.; Wu, Y.; Yang, Y.; Tso, P.; Wu, Z. Interactions between Intestinal Microbiota and Host Immune Response in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 942. [Google Scholar] [CrossRef]
- Wright, E.K.; Ding, N.S.; Niewiadomski, O. Management of Inflammatory Bowel Disease. Med. J. Aust. 2018, 209, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Quezada, S.M.; McLean, L.P.; Cross, R.K. Adverse events in IBD therapy: The 2018 update. Expert. Rev. Gastroenterol. Hepatol. 2018, 12, 1183–1191. [Google Scholar] [CrossRef]
- Zenlea, T.; Peppercorn, M.A. Immunosuppressive therapies for inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 3146–3152. [Google Scholar] [CrossRef] [PubMed]
- Seyedian, S.S.; Nokhostin, F.; Malamir, M.D. A review of the diagnosis, prevention, and treatment methods of inflammatory bowel disease. J. Med. Life 2019, 12, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Zurba, Y.; Gros, B.; Shehab, M. Exploring the Pipeline of Novel Therapies for Inflammatory Bowel Disease; State of the Art Review. Biomedicines 2023, 11, 747. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J.; Panés, J.; Danese, S.; Sharafali, Z.; Hassanali, A.; Jacob-Moffatt, R.; Eden, C.; Daperno, M.; Valentine, J.F.; Laharie, D.; et al. Etrolizumab as induction and maintenance therapy in patients with moderately to severely active Crohn’s disease (BERGAMOT): A randomised, placebo-controlled, double-blind, phase 3 trial. Lancet Gastroenterol. Hepatol. 2023, 8, 43–55. [Google Scholar] [CrossRef]
- Qiu, Y.; Chen, B.L.; Mao, R.; Zhang, S.H.; He, Y.; Zeng, Z.R.; Ben-Horin, S.; Chen, M.H. Systematic review with meta-analysis: Loss of response and requirement of anti-TNFα dose intensification in Crohn’s disease. J. Gastroenterol. 2017, 52, 535–554. [Google Scholar] [CrossRef]
- Katsanos, K.H.; Papamichael, K.; Feuerstein, J.D.; Christodoulou, D.K.; Cheifetz, A.S. Biological therapies in inflammatory bowel disease: Beyond anti-TNF therapies. Clin. Immunol. 2019, 206, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Adegbola, S.O.; Sahnan, K.; Warusavitarne, J.; Hart, A.; Tozer, P. Anti-TNF Therapy in Crohn’s Disease. Int. J. Mol. Sci. 2018, 19, 2244. [Google Scholar] [CrossRef] [PubMed]
- Sabino, J.; Verstockt, B.; Vermeire, S.; Ferrante, M. New biologics and small molecules in inflammatory bowel disease: An update. Ther. Adv. Gastroenterol. 2019, 12, 1756284819853208. [Google Scholar] [CrossRef] [PubMed]
- Salzmann, M.; von Graffenried, T.; Righini-Grunder, F.; Braegger, C.; Spalinger, J.; Schibli, S.; Schoepfer, A.; Nydegger, A.; Pittet, V.; Sokollik, C.; et al. Drug-Related Adverse Events Necessitating Treatment Discontinuation in Pediatric Inflammatory Bowel Disease Patients. J. Pediatr. Gastroenterol. Nutr. 2022, 75, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Kang, E.A.; Park, S.K.; Hong, S.N.; Kim, Y.S.; Bang, K.B.; Kim, K.O.; Lee, H.S.; Kang, S.B.; Shin, S.Y.; et al. Long-term Outcomes after the Discontinuation of Anti-Tumor Necrosis Factor-α Therapy in Patients with Inflammatory Bowel Disease under Clinical Remission: A Korean Association for the Study of Intestinal Disease Multicenter Study. Gut Liver 2021, 15, 752–762. [Google Scholar] [CrossRef]
- Wong, D.J.; Roth, E.M.; Feuerstein, J.D.; Poylin, V.Y. Surgery in the age of biologics. Gastroenterol. Report. 2019, 7, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.; Frontali, A.; Baldi, C.; Cigognini, M.; Lamperti, G.; Manzo, C.A.; Maconi, G.; Ardizzone, S.; Foschi, D.; Sampietro, G.M. Repeated surgery for recurrent Crohn’s disease: Does the outcome keep worsening operation after operation? A comparative study of 1224 consecutive procedures. Updates Surg. 2022, 74, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Moreno, L.O.; Fernández-Tomé, S.; Abalo, R. Biological Treatments in Inflammatory Bowel Disease: A Complex Mix of Mechanisms and Actions. Biologics 2021, 1, 189–210. [Google Scholar] [CrossRef]
- Lichtenstein, G.R.; Shahabi, A.; Seabury, S.A.; Lakdawalla, D.N.; Espinosa, O.D.; Green, S.; Brauer, M.; Baldassano, R.N. Lifetime Economic Burden of Crohn’s Disease and Ulcerative Colitis by Age at Diagnosis. Clin. Gastroenterol. Hepatol. 2020, 18, 889–897.e10. [Google Scholar] [CrossRef]
- Jannas-Vela, S.; Espinosa, A.; Candia, A.A.; Flores-Opazo, M.; Peñailillo, L.; Valenzuela, R. The Role of Omega-3 Polyunsaturated Fatty Acids and Their Lipid Mediators on Skeletal Muscle Regeneration: A Narrative Review. Nutrients 2023, 15, 871. [Google Scholar] [CrossRef]
- Van Dael, P. Role of n-3 long-chain polyunsaturated fatty acids in human nutrition and health: Review of recent studies and recommendations. Nutr. Res. Pract. 2021, 15, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- MacLean, C.H.; Mojica, W.A.; Newberry, S.J.; Pencharz, J.; Garland, R.H.; Tu, W.; Hilton, L.G.; Gralnek, I.M.; Rhodes, S.; Khanna, P.; et al. Systematic review of the effects of n-3 fatty acids in inflammatory bowel disease. Am. J. Clin. Nutr. 2005, 82, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Mozaffari, H.; Daneshzad, E.; Larijani, B.; Bellissimo, N.; Azadbakht, L. Dietary intake of fish, n-3 polyunsaturated fatty acids, and risk of inflammatory bowel disease: A systematic review and meta-analysis of observational studies. Eur. J. Nutr. 2020, 59, 1–17. [Google Scholar] [CrossRef]
- Zhang, T.; Li, G.; Duan, M.; Lv, T.; Feng, D.; Lu, N.; Zhou, Y.; Gu, L.; Zhu, W.; Gong, J. Perioperative parenteral fish oil supplementation improves postoperative coagulation function and outcomes in patients undergoing colectomy for ulcerative colitis. JPEN J. Parenter. Enter. Nutr. 2022, 46, 878–886. [Google Scholar] [CrossRef]
- Fu, J.; Chai, C.; Chen, L.; Cai, M.; Ai, B.; Li, H.; Yuan, J.; Lin, H.; Zhang, Z. Associations of Fish and Fish Oil Consumption with Incident Inflammatory Bowel Disease: A Population-Based Prospective Cohort Study. Inflamm. Bowel Dis. 2023; Online ahead of print. [Google Scholar]
- Klek, S.; Mankowska-Wierzbicka, D.; Scislo, L.; Walewska, E.; Pietka, M.; Szczepanek, K. High Dose Intravenous Fish Oil Reduces Inflammation-A Retrospective Tale from Two Centers. Nutrients 2020, 12, 2865. [Google Scholar] [CrossRef]
- Fu, Y. Marine n-3 polyunsaturated fatty acids and inflammatory diseases. In Advances in Dietary Lipids and Human Health; Elsevier: Amsterdam, The Netherlands, 2022; pp. 225–242. [Google Scholar]
- Fuller, I.D.; Cumming, A.H.; Card, A.; Burgess, E.J.; Barrow, C.J.; Perry, N.B.; Killeen, D.P. Free fatty acids in commercial krill oils: Concentrations, compositions, and implications for oxidative stability. J. Am. Oil Chem. Soc. 2020, 97, 889–900. [Google Scholar] [CrossRef]
- Xie, D.; Gong, M.; Wei, W.; Jin, J.; Wang, X.; Wang, X.; Jin, Q. Antarctic Krill (Euphausia superba) Oil: A Comprehensive Review of Chemical Composition, Extraction Technologies, Health Benefits, and Current Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 514–534. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, A.G.; Hassanzadeganroudsari, M.; Jovanovska, V.; Luwor, R.B.; Nurgali, K.; Su, X.Q. The comparative anti-cancer effects of krill oil and oxaliplatin in an orthotopic mouse model of colorectal cancer. Nutr. Metab. 2022, 19, 12. [Google Scholar] [CrossRef]
- Jayathilake, A.G.; Senior, P.V.; Su, X.Q. Krill oil extract suppresses cell growth and induces apoptosis of human colorectal cancer cells. BMC Complement. Altern. Med. 2016, 16, 328. [Google Scholar] [CrossRef]
- Jayathilake, A.G.; Kadife, E.; Luwor, R.B.; Nurgali, K.; Su, X.Q. Krill oil extract suppresses the proliferation of colorectal cancer cells through activation of caspase 3/9. Nutr. Metab. 2019, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Murzina, S.A.; Voronin, V.P.; Bitiutskii, D.G.; Mishin, A.V.; Khurtina, S.N.; Frey, D.I.; Orlov, A.M. Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat. J. Mar. Sci. Eng. 2023, 11, 1912. [Google Scholar] [CrossRef]
- Colletti, A.; Cravotto, G.; Citi, V.; Martelli, A.; Testai, L.; Cicero, A.F.G. Advances in Technologies for Highly Active Omega-3 Fatty Acids from Krill Oil: Clinical Applications. Mar. Drugs 2021, 19, 306. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liao, D.; He, B.; Zhou, G.; Cui, Y. Clinical effectiveness of krill oil supplementation on cardiovascular health in humans: An updated systematic review and meta-analysis of randomized controlled trials. Diabetes Metab. Syndr. Clin. Res. Rev. 2023, 17, 102909. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.G.; El-Kashef, D.H. Krill oil alleviates oxidative stress, iron accumulation and fibrosis in the liver and spleen of iron-overload rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 3950–3961. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Jiang, M.; Huang, W.; Liu, S. Antarctic Krill Oil Attenuates Oxidative Stress via the KEAP1-NRF2 Signaling in Patients with Coronary Heart Disease. Evid. Based Complement. Altern. Med. 2020, 2020, 9534137. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, A.G.; Kadife, E.; Kuol, N.; Luwor, R.B.; Nurgali, K.; Su, X.Q. Krill oil supplementation reduces the growth of CT-26 orthotopic tumours in Balb/c mice. BMC Complement. Med. Ther. 2022, 22, 34. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Lim, S.J.; Ryu, Y.M.; Park, H.R.; Suh, H.J.; Han, S.H. Absorption rate of krill oil and fish oil in blood and brain of rats. Lipids Health Dis. 2018, 17, 162. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, J.F.; Harvey, L.; Aziz, S.; Jordan, H.; Hegele, R.A.; Lemieux, P. A Single-dose, Comparative Bioavailability Study of a Formulation containing OM3 as Phospholipid and Free Fatty Acid to an Ethyl Ester Formulation in the Fasting and Fed States. Clin. Ther. 2019, 41, 426–444. [Google Scholar] [CrossRef]
- Sung, H.H.; Sinclair, A.J.; Lewandowski, P.A.; Su, X.Q. Postprandial long-chain n-3 polyunsaturated fatty acid response to krill oil and fish oil consumption in healthy women: A randomised controlled, single-dose, crossover study. Asia Pac. J. Clin. Nutr. 2018, 27, 148–157. [Google Scholar]
- Stolfi, C.; Maresca, C.; Monteleone, G.; Laudisi, F. Implication of Intestinal Barrier Dysfunction in Gut Dysbiosis and Diseases. Biomedicines 2022, 10, 289. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert. Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Slifer, Z.M.; Blikslager, A.T. The Integral Role of Tight Junction Proteins in the Repair of Injured Intestinal Epithelium. Int. J. Mol. Sci. 2020, 21, 972. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Wang, Y.; He, K.; Zhu, S. Microbial sensing in the intestine. Protein Cell 2023, 14, 824–860. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal. Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef]
- Li, Q.; von Ehrlich-Treuenstätt, V.; Schardey, J.; Wirth, U.; Zimmermann, P.; Andrassy, J.; Bazhin, A.V.; Werner, J.; Kühn, F. Gut Barrier Dysfunction and Bacterial Lipopolysaccharides in Colorectal Cancer. J. Gastrointest. Surg. 2023, 27, 1466–1472. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cui, W.; Li, X.; Yang, H. Interaction Between Commensal Bacteria, Immune Response and the Intestinal Barrier in Inflammatory Bowel Disease. Front. Immunol. 2021, 12, 761981. [Google Scholar] [CrossRef]
- Chang, J.T. Pathophysiology of Inflammatory Bowel Diseases. N. Engl. J. Med. 2020, 383, 2652–2664. [Google Scholar] [CrossRef]
- Costanzo, M.; Cesi, V.; Prete, E.; Negroni, A.; Palone, F.; Cucchiara, S.; Oliva, S.; Leter, B.; Stronati, L. Krill oil reduces intestinal inflammation by improving epithelial integrity and impairing adherent-invasive Escherichia coli pathogenicity. Dig. Liver Dis. 2016, 48, 34–42. [Google Scholar] [CrossRef]
- Liu, F.; Smith, A.D.; Solano-Aguilar, G.; Wang, T.T.; Pham, Q.; Beshah, E.; Tang, Q.; Urban, J.F.; Xue, C.; Li, R.W. Mechanistic insights into the attenuation of intestinal inflammation and modulation of the gut microbiome by krill oil using in vitro and in vivo models. Microbiome 2020, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xiang, X.; Zhou, Y.; Zhou, T.; Deng, S.; Zheng, B.; Zheng, P. Protective effects of Antarctic krill oil in dextran sulfate sodium-induced ulcerative colitis mice. J. Funct. Foods 2021, 79, 104394. [Google Scholar] [CrossRef]
- Kim, J.H.; Hong, S.S.; Lee, M.; Lee, E.H.; Rhee, I.; Chang, S.Y.; Lim, S.J. Krill Oil-Incorporated Liposomes as An Effective Nanovehicle to Ameliorate the Inflammatory Responses of DSS-Induced Colitis. Int. J. Nanomed. 2019, 14, 8305–8320. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Smith, A.D.; Wang, T.T.; Pham, Q.; Hou, P.; Cheung, L.; Yang, H.; Li, R.W. Phospholipid-rich krill oil promotes intestinal health by strengthening beneficial gut microbial interactions in an infectious colitis model. Food Funct. 2024, 15, 2604–2615. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Li, Y.Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Brand, S. Crohn’s disease: Th1, Th17 or both? The change of a paradigm: New immunological and genetic insights implicate Th17 cells in the pathogenesis of Crohn’s disease. Gut 2009, 58, 1152–1167. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.A.; Boucher, G.; Lees, C.W.; Franke, A.; D’Amato, M.; Taylor, K.D.; Lee, J.C.; Goyette, P.; Imielinski, M.; Latiano, A.; et al. Meta-analysis identifies 29 additional ulcerative colitis risk loci, increasing the number of confirmed associations to 47. Nat. Genet. 2011, 43, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Tilg, H.; Raine, T. IL-12, IL-23 and IL-17 in IBD: Immunobiology and therapeutic targeting. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Hamza, T.; Barnett, J.B.; Li, B. Interleukin 12 a Key Immunoregulatory Cytokine in Infection Applications. Int. J. Mol. Sci. 2010, 11, 789–806. [Google Scholar] [CrossRef]
- Álvarez-Salamero, C.; Castillo-González, R.; Pastor-Fernández, G.; Mariblanca, I.R.; Pino, J.; Cibrian, D.; Navarro, M.N. IL-23 signaling regulation of pro-inflammatory T-cell migration uncovered by phosphoproteomics. PLoS Biol. 2020, 18, e3000646. [Google Scholar] [CrossRef]
- Pedersen, J.; Coskun, M.; Soendergaard, C.; Salem, M.; Nielsen, O.H. Inflammatory pathways of importance for management of inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Jeen, Y.T. Anti-integrin therapy for inflammatory bowel disease. World J. Gastroenterol. 2018, 24, 1868–1880. [Google Scholar] [CrossRef] [PubMed]
- Paramsothy, S.; Rosenstein, A.K.; Mehandru, S.; Colombel, J.F. The current state of the art for biological therapies and new small molecules in inflammatory bowel disease. Mucosal Immunol. 2018, 11, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Hemperly, A.; Vande Casteele, N. Clinical Pharmacokinetics and Pharmacodynamics of Infliximab in the Treatment of Inflammatory Bowel Disease. Clin. Pharmacokinet. 2018, 57, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Grimstad, T.; Bjørndal, B.; Cacabelos, D.; Aasprong, O.G.; Janssen, E.A.; Omdal, R.; Svardal, A.; Hausken, T.; Bohov, P.; Portero-Otin, M.; et al. Dietary supplementation of krill oil attenuates inflammation and oxidative stress in experimental ulcerative colitis in rats. Scand. J. Gastroenterol. 2012, 47, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Yao, J.; Zhou, L.; Li, X.; Zhu, J.; Hu, Y.; Liu, J. Protective effect and mechanism insight of purified Antarctic kill phospholipids against mice ulcerative colitis combined with bioinformatics. Nat. Prod. Bioprospect 2023, 13, 11. [Google Scholar]
- Bonaterra, G.A.; Driscoll, D.; Schwarzbach, H.; Kinscherf, R. Krill Oil-In-Water Emulsion Protects against Lipopolysaccharide-Induced Proinflammatory Activation of Macrophages In Vitro. Mar. Drugs 2017, 15, 74. [Google Scholar] [CrossRef]
- Park, D.R.; Ko, R.; Kwon, S.H.; Min, B.; Yun, S.H.; Kim, M.H.; Minatelli, J.; Hill, S.; Lee, S.Y. FlexPro MD, a Mixture of Krill Oil, Astaxanthin, and Hyaluronic Acid, Suppresses Lipopolysaccharide-Induced Inflammatory Cytokine Production Through Inhibition of NF-κB. J. Med. Food 2016, 19, 1196–1203. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Shan, J.-L.; He, H.-T.; Li, M.-X.; Zhu, J.-W.; Cheng, Y.; Hu, N.; Wang, G.; Wang, D.; Yang, X.-Q.; He, Y.; et al. APE1 promotes antioxidant capacity by regulating Nrf-2 function through a redox-dependent mechanism. Free Radic. Biol. Med. 2015, 78, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Lih-Brody, L.; Powell, S.R.; Collier, K.P.; Reddy, G.M.; Cerchia, R.; Kahn, E.; Weissman, G.S.; Katz, S.; Floyd, R.A.; McKinley, M.J.; et al. Increased oxidative stress and decreased antioxidant defenses in mucosa of inflammatory bowel disease. Dig. Dis. Sci. 1996, 41, 2078–2086. [Google Scholar] [CrossRef] [PubMed]
- Hofseth, L.J.; Khan, M.A.; Ambrose, M.; Nikolayeva, O.; Xu-Welliver, M.; Kartalou, M.; Hussain, S.P.; Roth, R.B.; Zhou, X.; Mechanic, L.E.; et al. The adaptive imbalance in base excision-repair enzymes generates microsatellite instability in chronic inflammation. J. Clin. Investig. 2003, 112, 1887–1894. [Google Scholar] [CrossRef]
- Chami, B.; Martin, N.J.J.; Dennis, J.M.; Witting, P.K. Myeloperoxidase in the inflamed colon: A novel target for treating inflammatory bowel disease. Arch. Biochem. Biophys. 2018, 645, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, X.; Zhang, S.; Qi, C.; Zhang, Z.; Ma, R.; Xiang, L.; Chen, L.; Zhu, Y.; Tang, C.; et al. Oxidative stress gene expression, DNA methylation, and gut microbiota interaction trigger Crohn’s disease: A multi-omics Mendelian randomization study. BMC Med. 2023, 21, 179. [Google Scholar] [CrossRef]
- Ordas, I.; Eckmann, L.; Talamini, M.; Baumgart, D.C.; Sandborn, W.J. Ulcerative colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of oxidative stress in inflammatory bowel disease and potential antioxidant therapies. Oxidative Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef]
- Oshitani, N.; Sawa, Y.; Hara, J.; Adachi, K.; Nakamura, S.; Matsumoto, T.; Arakawa, T.; Kuroki, T. Functional and phenotypical activation of leucocytes in inflamed human colonic mucosa. J. Gastroenterol. Hepatol. 1997, 12, 809–814. [Google Scholar] [CrossRef]
- Sahakian, L.; Robinson, A.M.; Sahakian, L.; Stavely, R.; Kelley, M.R.; Nurgali, K. APE1/Ref-1 as a Therapeutic Target for Inflammatory Bowel Disease. Biomolecules 2023, 13, 1569. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front. Endocrinol. 2023, 14, 1217165. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.; Huang, M.T.; Kwon, K.H.; Chan, J.Y.; Reddy, B.S.; Kong, A.N. Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 2006, 66, 11580–11584. [Google Scholar] [CrossRef] [PubMed]
- Gerstgrasser, A.; Melhem, H.; Leonardi, I.; Atrott, K.; Schäfer, M.; Werner, S.; Rogler, G.; Frey-Wagner, I. Cell-specific Activation of the Nrf2 Antioxidant Pathway Increases Mucosal Inflammation in Acute but Not in Chronic Colitis. J. Crohns Colitis 2017, 11, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; Kloska, D.; Grochot-Przęczek, A.; Feelisch, M.; Cuadrado, A.; van Goor, H. Personalized redox medicine in inflammatory bowel diseases: An emerging role for HIF-1α and NRF2 as therapeutic targets. Redox Biol. 2023, 60, 102603. [Google Scholar] [CrossRef] [PubMed]
- Fasano, E.; Serini, S.; Mondella, N.; Trombino, S.; Celleno, L.; Lanza, P.; Cittadini, A.; Calviello, G. Antioxidant and anti-inflammatory effects of selected natural compounds contained in a dietary supplement on two human immortalized keratinocyte lines. BioMed Res. Int. 2014, 2014, 327452. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jang, J.S.; Son, D.J.; Im, H.S.; Kim, J.Y.; Park, J.E.; Choi, W.R.; Han, S.B.; Hong, J.T. Antarctic Krill Oil Diet Protects against Lipopolysaccharide-Induced Oxidative Stress, Neuroinflammation and Cognitive Impairment. Int. J. Mol. Sci. 2017, 18, 2554. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.; Matsui, T. Advanced glycation end products, oxidative stress and diabetic nephropathy. Oxid. Med. Cell Longev. 2010, 3, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Jin, L.; Liu, C.; Zhang, R.; Siebert, H.-C.; Li, Y.; Loers, G.; Petridis, A.K.; Xia, Z.; Dong, H.; et al. An antarctic krill oil-based diet elicits neuroprotective effects by inhibiting oxidative stress and rebalancing the M1/M2 microglia phenotype in a cuprizone model for demyelination. J. Funct. Foods 2021, 76, 104309. [Google Scholar] [CrossRef]
- Xiong, Q.; Ru, Q.; Tian, X.; Zhou, M.; Chen, L.; Li, Y.; Li, C. Krill oil protects PC12 cells against methamphetamine-induced neurotoxicity by inhibiting apoptotic response and oxidative stress. Nutr. Res. 2018, 58, 84–94. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Ng, S.C. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef]
- Xu, X.; Ocansey, D.K.W.; Hang, S.; Wang, B.; Amoah, S.; Yi, C.; Zhang, X.; Liu, L.; Mao, F. The gut metagenomics and metabolomics signature in patients with inflammatory bowel disease. Gut Pathog. 2022, 14, 26. [Google Scholar] [CrossRef]
- Kang, D.Y.; Park, J.L.; Yeo, M.K.; Kang, S.B.; Kim, J.M.; Kim, J.S.; Kim, S.Y. Diagnosis of Crohn’s disease and ulcerative colitis using the microbiome. BMC Microbiol. 2023, 23, 336. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef]
- Shin, Y.; Han, S.; Kwon, J.; Ju, S.; Choi, T.G.; Kang, I.; Kim, S.S. Roles of Short-Chain Fatty Acids in Inflammatory Bowel Disease. Nutrients 2023, 15, 4466. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Li, T.; Li, M.; Huang, S.; Qiu, Y.; Feng, R.; Zhang, S.; Chen, M.; Xiong, L.; Zeng, Z. Systematic review and meta-analysis: Short-chain fatty acid characterization in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2019, 25, 1751–1763. [Google Scholar] [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A.J.; Yao, S.; Xiao, Y.; Huang, X.; Eaves-Pyles, T.D.; Golovko, G.; et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol. 2018, 11, 752–762. [Google Scholar] [CrossRef]
- Wechsler, J.B.; Szabo, A.; Hsu, C.L.; Krier-Burris, R.A.; Schroeder, H.A.; Wang, M.Y.; Carter, R.G.; Velez, T.E.; Aguiniga, L.M.; Brown, J.B.; et al. Histamine drives severity of innate inflammation via histamine 4 receptor in murine experimental colitis. Mucosal Immunol. 2018, 11, 861–870. [Google Scholar] [CrossRef]
- Heazlewood, C.K.; Cook, M.C.; Eri, R.; Price, G.R.; Tauro, S.B.; Taupin, D.; Thornton, D.J.; Png, C.W.; Crockford, T.L.; Cornall, R.J.; et al. Aberrant mucin assembly in mice causes endoplasmic reticulum stress and spontaneous inflammation resembling ulcerative colitis. PLoS Med. 2008, 5, e54. [Google Scholar] [CrossRef]
- Robinson, A.M.; Gondalia, S.; Karpe, A.; Eri, R.; Beale, D.; Morrison, P.; Palombo, E.; Nurgali, K. Fecal microbiota and metabolome in a mouse model of spontaneous chronic colitis: Relevance to human inflammatory bowel disease. Inflamm. Bowel Dis. 2016, 22, 2767–2787. [Google Scholar] [CrossRef]
- Rahman, A.A.; Robinson, A.M.; Jovanovska, V.; Eri, R.; Nurgali, K. Alterations in the distal colon innervation in Winnie mouse model of spontaneous chronic colitis. Cell Tissue Res. 2015, 362, 497–512. [Google Scholar] [CrossRef]
- Farrukh, A.; Mayberry, J.F. Is there a role for fish oil in inflammatory bowel disease? World J. Clin. Cases 2014, 2, 250–252. [Google Scholar] [CrossRef]
- Turner, D.; Shah, P.S.; Steinhart, A.H.; Zlotkin, S.; Griffiths, A.M. Maintenance of remission in inflammatory bowel disease using omega-3 fatty acids (fish oil): A systematic review and meta-analyses. Inflamm. Bowel Dis. 2011, 17, 336–345. [Google Scholar] [CrossRef]
- Huang, X.; Li, Y.; Zhuang, P.; Liu, X.; Zhang, Y.; Zhang, P.; Jiao, J. Habitual Fish Oil Supplementation and Risk of Incident Inflammatory Bowel Diseases: A Prospective Population-Based Study. Front. Nutr. 2022, 9, 905162. [Google Scholar] [CrossRef]
- Ajabnoor, S.M.; Thorpe, G.; Abdelhamid, A.; Hooper, L. Long-term effects of increasing omega-3, omega-6 and total polyunsaturated fats on inflammatory bowel disease and markers of inflammation: A systematic review and meta-analysis of randomized controlled trials. Eur. J. Nutr. 2021, 60, 2293–2316. [Google Scholar] [CrossRef]
- Seidner, D.L.; Lashner, B.A.; Brzezinski, A.; Banks, P.L.; Goldblum, J.; Fiocchi, C.; Katz, J.; Lichtenstein, G.R.; Anton, P.A.; Kam, L.Y. An oral supplement enriched with fish oil, soluble fiber, and antioxidants for corticosteroid sparing in ulcerative colitis: A randomized, controlled trial. Clin. Gastroenterol. Hepatol. 2005, 3, 358–369. [Google Scholar] [CrossRef]
- Romano, C.; Cucchiara, S.; Barabino, A.; Annese, V.; Sferlazzas, C.; SIGENP Italian Study Group of Pediatric Inflammatory Bowel Diseases. Usefulness of ω-3 fatty acid supplementation in addition to mesalazine in maintaining remission in pediatric Crohn’s disease: A double-blind, randomized, placebo-controlled study. World J. Gastroenterol. 2005, 11, 7118. [Google Scholar] [CrossRef]
- Chang, J.P.; Tseng, P.T.; Zeng, B.S.; Chang, C.H.; Su, H.; Chou, P.H.; Su, K.P. Safety of Supplementation of Omega-3 Polyunsaturated Fatty Acids: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2023, 14, 1326–1336. [Google Scholar] [CrossRef]
- Scaioli, E.; Sartini, A.; Bellanova, M.; Campieri, M.; Festi, D.; Bazzoli, F.; Belluzzi, A. Eicosapentaenoic acid reduces fecal levels of calprotectin and prevents relapse in patients with ulcerative colitis. Clin. Gastroenterol. Hepatol. 2018, 16, 1268–1275.e2. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Lincoff, A.M.; Garcia, M.; Bash, D.; Ballantyne, C.M.; Barter, P.J.; Davidson, M.H.; Kastelein, J.J.P.; Koenig, W.; McGuire, D.K.; et al. Effect of High-Dose Omega-3 Fatty Acids vs Corn Oil on Major Adverse Cardiovascular Events in Patients at High Cardiovascular Risk: The STRENGTH Randomized Clinical Trial. JAMA 2020, 324, 2268–2280. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the Tolerable Upper Intake Level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). EFSA J. 2012, 10, 2815. [Google Scholar]
- Miyoshi, J.; Matsuoka, K.; Yoshida, A.; Naganuma, M.; Hisamatsu, T.; Yajima, T.; Inoue, N.; Okamoto, S.; Iwao, Y.; Ogata, H.; et al. 5-Aminosalicylic acid aggravates colitis mimicking exacerbation of ulcerative colitis. Intest. Res. 2018, 16, 635–640. [Google Scholar] [CrossRef]
- Barrett, K.; Saxena, S.; Pollok, R. Using corticosteroids appropriately in inflammatory bowel disease: A guide for primary care. Br. J. Gen. Pract. 2018, 68, 497–498. [Google Scholar] [CrossRef]
- Harlan, W.R.; Meyer, A.; Fisher, J. Inflammatory Bowel Disease: Epidemiology, Evaluation, Treatment, and Health Maintenance. N. C. Med. J. 2016, 77, 198–201. [Google Scholar] [CrossRef]
- Bernstein, C.N. Treatment of IBD: Where we are and where we are going. Am. J. Gastroenterol. 2015, 110, 114–126. [Google Scholar] [CrossRef]
- Miyatani, Y.; Kobayashi, T. Evidence-Based Approach to the Discontinuation of Immunomodulators or Biologics in Inflammatory Bowel Disease. Digestion 2023, 104, 66–73. [Google Scholar] [CrossRef]
- Liu, J.; Di, B.; Xu, L.L. Recent advances in the treatment of IBD: Targets, mechanisms and related therapies. Cytokine Growth Factor. Rev. 2023, 71–72, 1–12. [Google Scholar]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef]
- Costanzo, M.; Cesi, V.; Palone, F.; Pierdomenico, M.; Colantoni, E.; Leter, B.; Vitali, R.; Negroni, A.; Cucchiara, S.; Stronati, L. Krill oil, vitamin D and Lactobacillus reuteri cooperate to reduce gut inflammation. Benef. Microbes 2018, 9, 389–399. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, J.; Zhou, L.; Kan, J.; Zhao, M.; Huang, R.; Liu, J.; Marchioni, E. Antarctic krill oil high internal phase Pickering emulsion stabilized by bamboo protein gels and the anti-inflammatory effect in vitro and in vivo. J. Funct. Foods 2022, 94, 105134. [Google Scholar] [CrossRef]
- Cho, J.Y.; Chi, S.G.; Chun, H.S. Oral administration of docosahexaenoic acid attenuates colitis induced by dextran sulfate sodium in mice. Mol. Nutr. Food Res. 2011, 55, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shi, P.; Sun, Y.; Sun, J.; Dong, J.-N.; Wang, H.-G.; Zuo, L.-G.; Gong, J.-F.; Li, Y.; Gu, L.-L. DHA protects against experimental colitis in IL-10-deficient mice associated with the modulation of intestinal epithelial barrier function. Br. J. Nutr. 2015, 114, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Liu, C.; Qin, Q.; Zhang, Y.; Wang, X.; Zhang, J.; Odle, J.; Lin, X.; Hu, C.A.; Liu, Y. EPA and DHA attenuate deoxynivalenol-induced intestinal porcine epithelial cell injury and protect barrier function integrity by inhibiting necroptosis signaling pathway. FASEB J. 2020, 34, 2483–2496. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Hao, Y.-M.; Yang, Y.-H.; Zheng, Y.; Wu, Z.-J.; Zhou, M.-Q.; Wang, B.-Z.; Wang, Y.-M.; Wu, H.; Su, G.-H. DHA-Enriched Phospholipids and EPA-Enriched Phospholipids Alleviate Lipopolysaccharide-Induced Intestinal Barrier Injury in Mice via a Sirtuin 1-Dependent Mechanism. J. Agric. Food Chem. 2022, 70, 2911–2922. [Google Scholar] [CrossRef] [PubMed]
- Escoula, Q.; Bellenger, S.; Narce, M.; Bellenger, J. Docosahexaenoic and Eicosapentaenoic Acids Prevent Altered-Muc2 Secretion Induced by Palmitic Acid by Alleviating Endoplasmic Reticulum Stress in LS174T Goblet Cells. Nutrients 2019, 11, 2179. [Google Scholar] [CrossRef] [PubMed]
- El Mahdy, R.N.; Nader, M.A.; Helal, M.G.; Abu-Risha, S.E.; Abdelmageed, M.E. Eicosapentaenoic acid mitigates ulcerative colitis-induced by acetic acid through modulation of NF-κB and TGF-β/EGFR signaling pathways. Life Sci. 2023, 327, 121820. [Google Scholar] [CrossRef] [PubMed]
- Prossomariti, A.; Scaioli, E.; Piazzi, G.; Fazio, C.; Bellanova, M.; Biagi, E.; Candela, M.; Brigidi, P.; Consolandi, C.; Balbi, T.; et al. Short-term treatment with eicosapentaenoic acid improves inflammation and affects colonic differentiation markers and microbiota in patients with ulcerative colitis. Sci. Rep. 2017, 7, 7458. [Google Scholar] [CrossRef]
- Che, H.; Li, H.; Song, L.; Dong, X.; Yang, X.; Zhang, T.; Wang, Y.; Xie, W. Orally Administered DHA-Enriched Phospholipids and DHA-Enriched Triglyceride Relieve Oxidative Stress, Improve Intestinal Barrier, Modulate Inflammatory Cytokine and Gut Microbiota, and Meliorate Inflammatory Responses in the Brain in Dextran Sodium Sulfate Induced Colitis in Mice. Mol. Nutr. Food Res. 2021, 65, e2000986. [Google Scholar]
- Henry, W.; Suparna, M.; Fiona, C.C.; Morag, T.; Henry, M.W.; Sarah, L.P.; Jade, A.S.; Phil, Q.; Giles, J.T.; Clare, L.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974. [Google Scholar]
- Weldon, S.M.; Mullen, A.C.; Loscher, C.E.; Hurley, L.A.; Roche, H.M. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of EPA and DHA are dependent upon time and dose-response elements associated with LPS stimulation in THP-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef]
- Honda, K.L.; Lamon-Fava, S.; Matthan, N.R.; Wu, D.; Lichtenstein, A.H. EPA and DHA exposure alters the inflammatory response but not the surface expression of Toll-like receptor 4 in macrophages. Lipids 2015, 50, 121–129. [Google Scholar] [CrossRef]
- Kikut, J.; Drozd, A.; Mokrzycka, M.; Grzybowska-Chlebowczyk, U.; Ziętek, M.; Szczuko, M. Are EPA and DHA Derivatives Involved in IBD Remission? J. Clin. Med. 2022, 11, 2388. [Google Scholar] [CrossRef]
- Lee, H.-N.; Choi, Y.-S.; Kim, S.H.; Zhong, X.; Kim, W.; Park, J.S.; Saeidi, S.; Han, B.W.; Kim, N.; Lee, H.S.; et al. Resolvin D1 suppresses inflammation-associated tumorigenesis in the colon by inhibiting IL-6-induced mitotic spindle abnormality. FASEB J. 2021, 35, e21432. [Google Scholar] [CrossRef]
- Takenouchi, S.; Imai, D.; Nakamura, T.; Murata, T. Efficient Attenuation of Dextran Sulfate Sodium-Induced Colitis by Oral Administration of 5,6-Dihydroxy-8Z, 11Z, 14Z, 17Z-eicosatetraenoic Acid in Mice. Int. J. Mol. Sci. 2021, 22, 9295. [Google Scholar] [CrossRef]
- Chan, S.; Luben, R.; Olsen, A.; Tjonneland, A.; Kaaks, R.; Lindgren, S.; Grip, O.; Bergmann, M.; Boeing, H.; Hallmans, G. Association between high dietary intake of the n-3 polyunsaturated fatty acid docosahexaenoic acid and reduced risk of Crohn’s disease. Aliment. Pharmacol. Ther. 2014, 39, 834–842. [Google Scholar] [CrossRef]
- Jia, X.; Hu, C.; Wu, X.; Qi, H.; Lin, L.; Xu, M.; Xu, Y.; Wang, T.; Zhao, Z.; Chen, Y.; et al. Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study. Metabolites 2023, 13, 1041. [Google Scholar] [CrossRef]
- Uchiyama, K.; Nakamura, M.; Odahara, S.; Koido, S.; Katahira, K.; Shiraishi, H.; Ohkusa, T.; Fujise, K.; Tajiri, H. N-3 polyunsaturated fatty acid diet therapy for patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2010, 16, 1696–1707. [Google Scholar] [CrossRef]
- Feagan, B.G.; Sandborn, W.J.; Mittmann, U.; Bar-Meir, S.; D’Haens, G.; Bradette, M.; Cohen, A.; Dallaire, C.; Ponich, T.P.; McDonald, J.W.; et al. Omega-3 free fatty acids for the maintenance of remission in Crohn disease: The EPIC Randomized Controlled Trials. JAMA 2008, 299, 1690–1697. [Google Scholar] [CrossRef] [PubMed]
- Lev-Tzion, R.; Griffiths, A.M.; Leder, O.; Turner, D. Omega 3 fatty acids (fish oil) for maintenance of remission in Crohn’s disease. Cochrane Database Syst. Rev. 2014, 2014, Cd006320. [Google Scholar] [CrossRef] [PubMed]
- Sabo, C.M.; Simiras, C.; Ismaiel, A.; Dumitrascu, D.L. Diet and Gut Inflammation: The Effect of Diet on Inflammatory Markers in Inflammatory Bowel Disease—A Scoping Review. J. Gastrointestin Liver Dis. 2023, 32, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Kwantes, J.M.; Grundmann, O. A brief review of krill oil history, research, and the commercial market. J. Diet. Suppl. 2015, 12, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Hosokawa, M.; Mikami, N.; Miyashita, K.; Tanaka, T. Dietary astaxanthin inhibits colitis and colitis-associated colon carcinogenesis in mice via modulation of the inflammatory cytokines. Chem.-Biol. Interact. 2011, 193, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, Y.; Wu, S.; Zheng, W.; Song, S.; Ai, C. Fabrication of astaxanthin-enriched colon-targeted alginate microspheres and its beneficial effect on dextran sulfate sodium-induced ulcerative colitis in mice. Int. J. Biol. Macromol. 2022, 205, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Nishida, A.; Ohno, M.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; Andoh, A. Astaxanthin, a xanthophyll carotenoid, prevents development of dextran sulphate sodium-induced murine colitis. J. Clin. Biochem. Nutr. 2019, 64, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Gao, Q.; Yang, L.; Wang, X.; Wang, Z.; Li, Z.; Xu, J.; Xue, C. In-Depth Analysis of the Mechanism of Astaxanthin Succinate Diester in Reducing Ulcerative Colitis in C57BL/6J Mice Based on Microbiota Informatics. Molecules 2023, 28, 6513. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Ndrepepa, G. Myeloperoxidase—A bridge linking inflammation and oxidative stress with cardiovascular disease. Clin. Chim. Acta 2019, 493, 36–51. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Robinson, A.M.; Su, X.Q.; Nurgali, K. Krill Oil and Its Bioactive Components as a Potential Therapy for Inflammatory Bowel Disease: Insights from In Vivo and In Vitro Studies. Biomolecules 2024, 14, 447. https://doi.org/10.3390/biom14040447
Liu Y, Robinson AM, Su XQ, Nurgali K. Krill Oil and Its Bioactive Components as a Potential Therapy for Inflammatory Bowel Disease: Insights from In Vivo and In Vitro Studies. Biomolecules. 2024; 14(4):447. https://doi.org/10.3390/biom14040447
Chicago/Turabian StyleLiu, Yingying, Ainsley M. Robinson, Xiao Qun Su, and Kulmira Nurgali. 2024. "Krill Oil and Its Bioactive Components as a Potential Therapy for Inflammatory Bowel Disease: Insights from In Vivo and In Vitro Studies" Biomolecules 14, no. 4: 447. https://doi.org/10.3390/biom14040447
APA StyleLiu, Y., Robinson, A. M., Su, X. Q., & Nurgali, K. (2024). Krill Oil and Its Bioactive Components as a Potential Therapy for Inflammatory Bowel Disease: Insights from In Vivo and In Vitro Studies. Biomolecules, 14(4), 447. https://doi.org/10.3390/biom14040447