1. Introduction
Endophytic capabilities of fungi forming orchid mycorrhizas were observed many times in different plant families [
1,
2,
3]. The majority of these orchid symbionts belongs into a wide group of fungi mostly from phylum Basidiomycota, few members being from Ascomycota [
4]. In Basidiomycota phylum, we can distinguish one paraphyletic subgroup generally called
Rhizoctonia-like (
Rh-like) fungi or “rhizoctonias”, which include members of the families Ceratobasidiaceae and Tulasnellaceae and order Sebacinales. The morphological definition of
Rh-like fungi includes monilioid cells, unimpaired parenthosomes getting into doliporus septa and right-angle branching of hyphae [
4,
5,
6]. The former methods of their identification were based mainly on morphological parameters, while recent identification of
Rh-like fungi is based on the molecular methodology as the sequencing of, e.g., ITS regions of rDNA [
7]. Genus
Rhizoctonia was established historically by de Candolle (1815) and is coherent to
Rhizoctonia solani, now this genus is justified as a paraphyletic group encompassing species from different clades. Their pathogenicity depends on the genotype of a fungal strain and host plant species [
8]. The fungi from
Rh-like group are cosmopolitans mostly with saprotrophic abilities [
4], but the life strategy and other ecological aspects of these endophytes are not well-known [
3].
Most of the research was done on the members of the order Sebacinales, especially on the genus
Serendipita. The effects and mutualistic ability (growth promotion) was observed mostly in
Serendipita indica (syn.:
Piriformospora indica),
Serendipita herbamans, and
Serendipita vermifera (syn.:
Sebacina vermifera) [
2,
7,
9,
10,
11]. These species also point to the potential in applied research or agriculture due to low host specificity [
3,
12,
13,
14]. The order Sebacinales is known to be involved in various mycorrhizal symbioses and endophytic associations. However, other fungi forming orchid mycorrhiza are also capable of initiation endophytic symbiosis [
3,
7,
15]. As the ability of many
Rh-like fungi to colonize roots of different plant species differ,
Rh-like fungi do affect the species structure of vegetation [
16]. The substrate competition and niche occupation may play a role in spreading of pathogen or disease outbreaking, but the disease protective effects of
Rh-like fungi may also encompass systemic resistance or systemically acquired resistance [
12,
17]. Isolates of
Rh-like fungi forming a mycorrhizal symbiosis with orchids are used due to their non-pathogenic action for biological protection of plants against their phylogenetically related representatives, which cause significant economic losses [
18,
19]. Orchid tissues contain a wide range of fungi that grow endophytically, which have protective effects against plant pathogens [
20].
With the current emphasis on the sustainability of food production and the reduction of the use of chemical plant protection products, the study of the properties of endophytic organisms in the context of biological protection and the ability to improve crops is a key issue. Biological plant protection preparations offer a variety of effects, but it presupposes knowledge of broader context at the level of the plant and of the whole community [
17,
21]. Usually, the effect of chemical protection is faster, more effective and in many cases the use of specific chemical substances is inevitable [
22,
23]. Chemical protection can also affect non-target organisms, the quality of life of the surrounding population, or cause a phytotoxic effect on plants [
24]. In contrast, biological protection through
Rh-like fungi can mediate the translocation of a nutrient pool or provide other beneficial features [
15,
25,
26,
27]. If biological control agents are to be used on a wider scale in agriculture, they should be able to reduce the incidence of major cereal pathogens such as
Fusarium species complex.
Fusarium species, especially
F. culmorum, are a group of fungal plant pathogens that are responsible for a decrease of yield and quality of grain production constantly every year with rarely epidemic duration [
28].
F. culmorum causes two diseases on wheat: Fusarium root rot and Fusarium head blight. Fusarium root rot symptoms are pre- and post-emergence seedling death, or brown discolouration on the coleoptiles, roots and the pseudostem [
28]. Fusarium head blight symptoms include partial head blighting, with the prematurely bleached spikelets, or blighting of the entire head [
28,
29].
F. culmorum is pathogen causing Fusarium head blight in most places in Europe and is considered to be one of the main pathogens of wheat worldwide [
30], which significantly reduces the yield and quality of harvested wheat and barley grains [
28]. Lower yield production is mainly caused by
Fusarium-damaged kernels or spikelet sterility [
31,
32,
33]. The quality of contaminated grains is also lower due to the reduced content of nutrients and due to the presence of mycotoxins, which are toxic for humans and animals. The costs of fungicides or treatment of affected animals are enormous. Additionally, the reduction in biomass and grain yield noticeably decreases income in the agriculture industry [
34,
35,
36]. The manifestations of the disease are influenced by many factors such as temperature, humidity, genetic constitution both of the host and the pathogen, biotic or abiotic stresses, preceding crop, rotation of crops or agrotechnical management [
17,
37,
38,
39]. These environmental factors play an important role in the management of the disease by fungicides or by biocontrol agents [
38,
40].
The world’s most serious pathogens can also include pathogenic representatives of
Rh-like fungi.
C. cereale as a teleomorph of
R. cerealis causing a sharp eye spot on the bases of cereals, especially wheat. Severe infection can result in the death of young tillers or cause lodging of stems [
41]. Infestation with this pathogen allows other fungi to colonize host tissues, include highly virulent pathogens represented by
F. culmorum, or other weaker or non-pathogenic species of fungi such as
Microdochium bolleyi [
41].
M. bolleyi can be considered as a potential biocontrol agent against aggressive soil-borne pathogens in cereal crops [
42].
The main objective of this work is to observe the effects of fungi forming a mycorrhizal symbiosis with orchids against cereal pathogens. In this study, three independent experiments were carried out. We analysed the competitive abilities of varia Rh-like fungi, specifically five isolates of orchid mycorrhizal fungi from the genera Tulasnella and Ceratobasidium, endophytic M. bolleyi, and pathogenic C. cereale on the growth of pathogenic fungi F. culmorum and Pyrenophora teres under in vitro conditions. We also tested the abilities of the same Rh-like fungi and a well-known beneficial endophyte S. indica to ameliorate Fusarium root rot of wheat caused by F. culmorum grown in a growth chamber and the introduction of these fungi into field conditions for qualitative and quantitative yield parameters and the occurrence of barley diseases. The effects of these microorganisms were compared with fungicidal treatment with standard seed dressing fungicide containing prochloraz and triticonazole (Kinto Duo).
3. Discussion
This work is a part of the research on endophytic interactions of a group of fungi generally called
Rh-like fungi. Representatives of this group can be found across continents in diverse ecosystems, where their functions remain unexplored [
7] and are studied mainly in relation to plant-pathogen interactions [
17] and relation to orchid mycorrhiza [
27,
43]. Testing of local species as potential biocontrol agents is becoming more and more important. Similar cosmopolitan organisms are arbuscular mycorrhizal fungi, which as a natural part of ecosystems represent a promising opportunity in sustainable agriculture and plant protection. However, their limitation is the difficulty of cultivation as they are obligatory biotrophs [
44,
45,
46,
47]. In contrast, the endophytic fungi used in the experiment as well as many of other
Rh-like fungi can be cultivated on a large scale on an organic substrate, which makes their potential use more efficient and cheaper.
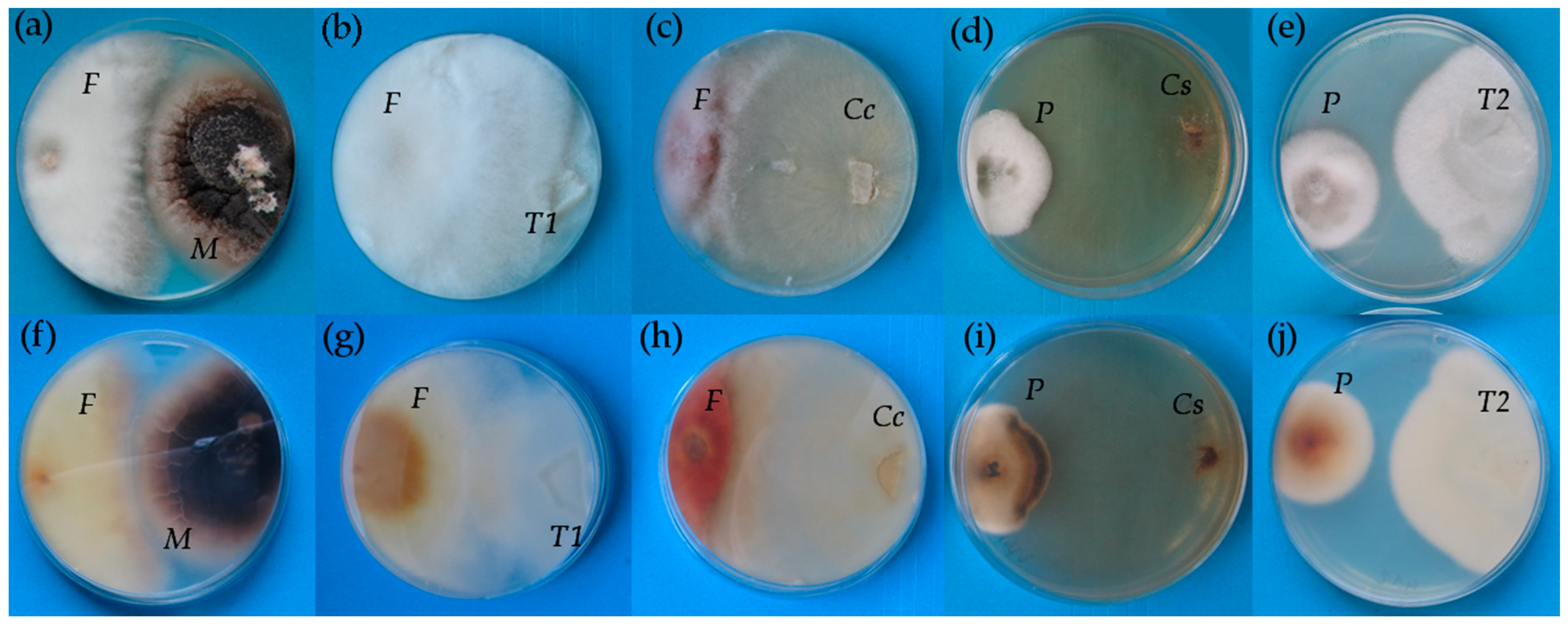
In vitro testing is commonly used as a first approach to assess traits of microorganisms studied [
48,
49]. Additionally, in our study, Petri dish tests were useful for comparison of
Rh-like fungi, which were isolated from orchid roots as mycorrhizal fungi with two other
Rh-like fungi as regard their abilities to compete against selected pathogens in competition.
C. cereale, which is generally considered as a plant pathogen [
50,
51], was in the current study the only isolate able to compete with both the pathogenic fungi used, i.e.,
F. culmorum and
P. teres, due to the action of metabolites, which uses as a pathogen to attack plants [
52]. Only two of five isolates from orchid roots were able to cause darkening of the
P. teres mycelia, which is a sign of degradation, but the reason for the action of only two isolates from this phylogenetically and ecologically related group of fungi remains unexplained and requires a more thorough analysis of metabolite interactions.
Although in vitro testing may indicate potentially efficient isolates of
Rh-like fungi, which we were able to confirm against
F. culmorum only in two isolates, in the case of
P. teres in three, so this proves the effectiveness of the isolates only partially, and it is necessary to test them under more realistic growth systems, especially with the target plants species. We used the pot experiments as a level suitable before the field application of obtained novel
Rh-like endophytes. Isolates whose effect against
F. culmorum was evident in in vitro tests did not show any protective effect in the pot experiments, besides the treatment of
C. cereale due to its pathogenic nature reduced the values of the evaluated parameters compared to the plants treated only with
F. culmorum. Our results stay in accordance with observations of Lemańczyk and Kwaśna [
41] who found that
C. cereale can reduce the resistance of plants to other pathogens. Biological protection of plants through these endophytes does not only have to take place through direct competition with the pathogen, but also by the induction of plant defence mechanisms [
17,
53]. The mechanism of induction of the defence reaction of our isolates is unknown, but the signalling particles may be fragments of cell walls during the formation of an endophytic association [
54].
The early stages of cereal development are well documented by the infectious pressure of fungi causing crown rot in the habitat [
28,
52]. The first and second pot experiments simulated the natural reproductive cycle of the
Fusarium spp., in which the pathogen is present in the infected grains or its infectious particles are in the substrate [
28]. According to the analyses described by Blanco et al. [
55], the infected grain is the direct source of the earliest infections and allows the fungus to attack the sprouts. This is because the infectious particles are located under layers of the seed testa, which is a common way of transmitting of pathogens [
55,
56]. As we suspect, in our studies it enabled the pathogen easier enters the plant tissues compared to endophytes provided in both mentioned experiments from the outside. The third experiment simulated the infectious pressure of a pathogen, for which post-harvest crop residues serve as a source of spread, so the main infectious pressure comes at the seedling stage [
57]. In this experiment, the infection could occur after the germination of plants and do not affect the early stages of an endophytic association. This delayed pathogen application did not affect the values of colonization despite our expectations, but it may have played a role in the plant development due to sudden stress compared to previous experiments, which led to a later harvest of Experiment 3 versus Experiment 2 to maintain the same growth stage of wheat. The colonization rate was reduced in the first pot experiment with an organic substrate compared to the other experiments. This reduction can also be explained by competition in the substrate with early colonizing saprotrophs, or by choosing a saprotrophic trophic strategy of endophytic strains caused by the availability of organic matter in the substrate.
Fusarium spp. had a devastating effect during germination and seedling stage of plant development, but the presence of the pathogen in the tissues of asymptomatic young plants could remain unmanifested [
58,
59], which is a weak point of the methodology used to evaluate plant infestation. Although molecular or biochemical analysis of the samples would be required for the presence of the pathogen, visual evaluation appears to be a suitable choice in these more than month-long experiments [
59], because during that time, some of the symptoms of the infection will show. On the opposite, the browning of bases does not have to be caused directly by the
Fusarium hyphae but as the plant response to its presence in the roots [
55], but other factors and pathogens can cause this symptom [
50,
60]. Typical representatives of crown rot are pathogenic strains of the genus
Rhizoctonia sp. or
Ceratobasidium sp. [
50,
60,
61] and predominantly in field conditions, it is difficult to determine the pathogen only by visual observation. The rate of infestation could be affected by airborne transmission caused by the increased availability of infectious particles forming on infested plants in a pot. The infected plants could serve as another source of inoculum because the major reservoirs of
Fusarium spp. are crop residues on the substrate surface [
57]. The infection process by
F. culmorum is influenced by temperature, humidity, carbon and nitrogen availability and the ability to produce mycotoxins that may confer higher pathogenicity by inhibiting the plant defence response [
28]. To understand the complexity of biological plant protection, it would be appropriate to study the impact of pathogens on plants, during their development.
P. teres is a serious pathogen of cereals causing the net blotch of barley negatively influencing yield and quality of grains leading to significant economic loses [
62,
63]. In our region, it is usually not necessary to perform artificial inoculations of
P. teres, because the natural incidence is mostly high [
64]. Conditions of the experimental year nevertheless caused the low occurrence of this pathogen. Similarly, another common pathogen
Cochliobolus sativus [
65] did not occur in our plots during the experiment. The evaluation of the presence and impact of pathogens on plants in the field is influenced, among other by factors as sowing procedures and the type of the preceding crop which, as a non-target host, can prevent the mass spread of infectious particles in the habitat by limiting the life cycle of the species-specific pathogen [
60]. Although there are many influences and stressors in field conditions, oilseed rape as preceding crop played a crucial role in the results we achieved. Nonsignificant results in yield could be also caused by an excess of mineral nutrients, which affected the spreading of endophytes in the roots, as it is approved for arbuscular mycorrhiza [
66]. Similar to yields, grain quality parameters could differ if colonization was higher or in stressful conditions, where the endophytic potential would play a more important role [
67,
68].
Application of biocontrol agents into the field is a key component of experiments [
12,
14,
48]. Due to the results of root colonization, the spreading of the mycelia overgrown substrate seems to be an acceptable way of application for
Rh-like fungi; however, it could be optimized to achieve colonization values comparable to pot experiments.







{kind=link}
{kind=link}
{kind=link}