1. Introduction
Triticum aestivum L. (bread wheat) is an essential grain worldwide, including Egypt. It provides humanity their protein requirements [
1,
2]. In developing countries, the requirement for wheat grains increased every year and is predicted to reach 60% by 2050 [
3]. The agriculture of most regions has experienced adverse effects from climate change, which constitutes the main cause of biotic and abiotic stresses. Climate change (drought, high temperatures, pests, floods, storm, and disease epidemics) greatly affects crop production globally [
4]. Generally, wheat plays a fateful role in global food security, especially in Egypt’s food economy, from production and consumption perspectives [
5].
Rusts and powdery mildew are wheat pathogens that cause a vital decrease in wheat production [
5]. In wheat, three species of wheat rust were found: leaf or brown rust (
Puccinia triticina), stem or black rust (
Puccinia graminis f. sp.
tritici) and stripe or yellow rust (
Puccinia striiformis f. sp.
tritici) [
6,
7]. Rusts are caused by Puccinia spp., considered to be the most significant wheat disease and representing a big concern for wheat breeders and farmers [
8]. It leads to tremendous strike yield losses annually to wheat crops worldwide [
9,
10].
The worldwide plant production is decreased by at least 10% because of diseases and pests [
11]. Among wheat rust diseases, the disease of leaf rust is still the number one widespread and destructive wheat disease, and it is the most important biotic stress that limited the productivity of bread wheat in Egypt and worldwide [
12]. It has caused significant losses in grain yield, which have reached 23% [
13], and the losses in epidemic seasons have reached up to 50% [
14]. Leaf rust has the capability of adapting to varied climatological conditions; therefore, it is one of the most devastating diseases of wheat worldwide [
15].
In general, wheat leaf rust occurs more frequently and spreads widely disease compared to the other two wheat rusts (stem and stripe rust) and is more prevalent globally [
16]. The pathogen is able to spread thousands of kilometers via wind [
9] and acclimate to diverse temperatures [
17]. It causes severe damage to yields worldwide due to decreases grain quality and quantity (yield production) by greater than 50% in susceptible varieties under the pathogens’ favorable environmental conditions [
18].
Fungicides are the most favored method by breeders to fight fungal diseases, but they are expensive and can harm the environment [
19]. Genetic resistance breeding is an efficient, economical and environmentally friendly way to relieve pathogen damage [
20], saving time and effort compared to traditional methods [
21].
Genetic resistance to leaf rust is generally classified into two forms: adult-plant resistance (APR) and seedling resistance (ASR) [
8,
22]. Seedling resistance (ASR) is monogenic, usually expressed at all growth stages, controlled by a single (major effect) gene, and hypersensitive. On the contrary, adult plant resistance (APR) is polygenic, typically best expressed in adult plants controlled by multiple (minor effect) genes, and non-hypersensitive, slow rusting [
23]. Additionally, it can distinguish the resistance to leaf rust into two types, qualitative conferred by single resistance genes and quantitative resistance, facilitated by multiple genes [
24].
One of the efficient methods used for leaf rust disease control is host-genetic resistance. It is an economical, environmentally safe approach and eliminates the use of synthetic fungicides. Race-specific genes for resistance allow enough protection against only a few pathotypes of the pathogen and interact according to the gene-for-gene theory [
25]. Additionally, this method aimed to obtain high-yielding varieties characterized by high levels of resistance to major diseases, especially wheat leaf rust [
22].
Recently, it has become possible to directly identify the
Lr genes using specific primers targeting the
Lr genes itself or by using linked molecular markers, such as simple sequence repeat (SSR), sequence-tagged site (STS), sequence-characterized amplified regions (SCAR) and cleaved amplified polymorphic sequences (CAPS) [
26]. Molecular markers, particularly PCR-based markers, proved their efficient ability to identify resistance genes in varieties and to combine them accurately toward selecting lines with suitable gene pool combinations [
27]. Thus, the introgression of the leaf rust resistance genes (
Lr genes) into wheat varieties using molecular markers (specific or linked) is the superior approach of their safeguarding in terms of environmental protection [
26].
Most leaf rust (
Lr) resistance genes are conferred as major, seedling, or race-specific and follow up the gene-for-gene theory [
28]; unfortunately, when new races of the pathogen appear, they lose their efficacy [
29]. Presently, there is a crucial need for breeding programs focusing on producing varieties with adult plant resistance (APR) [
19]. Likewise, wheat varieties differ in disease resistance depending on which
Lr genes they carry, or, in other words, their
Lr gene pool [
26].
Nowadays, more than 100 leaf rust resistance (
Lr) genes/alleles have been characterized, recognized, and described in seedling and adult plant resistance to leaf rust in wheat and its relatives [
7,
30]. The majority of them are originated from hexaploid bread wheat or wild grass species related to wheat, whereas a limited number have been identified and characterized in tetraploid durum wheat [
31]. A few race non-specific
Lr genes, including
Lr34 and
Lr67, have been found, mainly at the adult stage, conferring resistance to multiple pathogen species [
32].
Unfortunately, limited information is known about the
Lr genes content within Egyptian wheat varieties [
33,
34]. Therefore, this study evaluates the disease resistance level of the 50 Egyptian wheat varieties against leaf rust at the adult plant stage under open field conditions (two successive seasons) and identifies varieties with slow-rusting characteristics. Additionally, we define the leaf rust resistance genes’ content in the studied Egyptian wheat varieties/collection to act as a core base for successful and effective breeding programs for leaf rust resistance.
2. Results
2.1. Field Evaluation of Leaf Rust Resistance
The tested wheat varieties were evaluated under field conditions for their final rust severity (FRS), area under the disease progress curve (AUDPC) and rate of leaf rust disease increase (r-value) traits. According to the obtained records during the two growing seasons and based on recorded values of the final rust severity (%), the tested varieties have been clustered under three main groups: (a) varieties with moderate to high levels of resistance (Mabrok, Sakha 95, Montana, Sohag 5, Misr 2 and Giza 168) (FRS ranged between 0.33 and 2.32%), (b) varieties with intermediate to low levels of resistance (Giza 139, Giza 144, Giza 155, Giza 156, Giza 157, Giza 160, Giza 167, Giza 171, Sakha 8, Sakha 61, Sakha 88, Sakha 92, Sakha 94, Sids 8, Sids 5, Sids 6, Sids 7, Gemmeiza 7, Sids 14, Romana, Hendy 62, Nubaria 1, Gemmeiza 5, Gemmeiza 3, Gemmeiza 11, Gemmeiza 12, BanySwif 1, BanySwif 5, BanySwif 6, BanySwif 7 and Sohag 4) (with FRS less than 30%), and (c) highly susceptible varieties (Gemmeiza 1, Gemmeiza 9, Giza162, Giza 163, Giza 164, Giza 165, Sids 1, Sids 2, Sids 3, Sakha 62, Sakha 69, Sohag 3 and BanySwif 4) (with FRS more than 60%); the data are presented in
Table 1.
Additionally, the area under disease progress curve (AUDPC) parameter we used as a convenient and reliable criterion to describe the performance of durable resistance in any wheat variety. It combines the infection period and the level of infection and uses this combination to estimate (AUDPC) values for the tested varieties. In general, the values of (AUDPC) were found to be higher in the second season than in the first season. Based on the AUDPC records of the two seasons, the tested varieties could be divided into the following three groups. The first group included wheat varieties with the lowest AUDPC (less than 100); these cultivars were designated as the resistance varieties or partial resistance. This group includes six wheat varieties (Mabrok, Sakha 95, Montana, Sohag 5, Misr 2 and Giza 168) and their AUDPC values ranged from 5 to 63. The second group included the varieties with intermediate and low AUDPC (ranged from 101 to 390), and these varieties revealed the lowest levels of adult plant resistance to leaf rust infection under field conditions. This group includes varieties Giza 139, Giza 144, Giza 155, Giza 156, Giza 157, Giza 160, Giza 167, Giza 171, Sakha 8, Sakha 61, Sakha 88, Sakha 92, Sakha 94, Sids 8, Sids 5, Sids 6, Sids 7, Gemmeiza 7, Sids 14, Romana, Hendy 62, Nubaria 1, Gemmeiza 5, Gemmeiza 3, Gemmeiza 11, Gemmeiza 12, BanySwif 1, BanySwif 5, BanySwif 6, BanySwif 7 and Sohag 4.
The third group included the wheat varieties that showed the highest estimated AUDPC levels; the varieties in this group were classified as the highly susceptible or fast leaf rusting Egyptian wheat varieties (Gemmeiza 1, Gemmeiza 9, Giza162, Giza 163, Giza 164, Giza 165, Sids 1, Sids 2, Sids 3, Sakha 62, Sakha 69, Sohag 3 and BanySwif 4) with AUDPC values ranged from 610 to 111.
Rusting progression in tested wheat varieties was estimated based on the “rate of leaf rust increase lower rates (r-value)” parameter, and the 50 varieties were classified into three categories based on their r-values: (a) varieties exhibited almost complete resistance (their r-value ranged from 0.030 to 0.080); (b) varieties with slow leaf rusting rates, which exhibited intermediate levels of r-value (ranged from 0.090 to 0.180); and (c) fast rusting or highly susceptible wheat varieties, where the r-values reached the maximum levels (ranged from 0.200 to 0.218).
The analysis of variance results showed that the mean squares of the 50 wheat genotypes were highly significant for all the studied disease parameters during the two growing seasons as shown in
Table 2. These results showed that all the genotypes differed in their response to leaf rust disease.
2.2. Screening for Leaf Rust Resistance Genes within Egyptian Wheat Collection
Fifty Egyptian wheat varieties were characterized using 28
Lr genes (
Lr1, Lr9, Lr10, Lr13, Lr19, Lr20, Lr21, Lr22a, Lr24, Lr25, Lr26, Lr27, Lr28, Lr29, Lr32, Lr34, Lr35, Lr36, Lr37, Lr39, Lr46, Lr47, Lr48, Lr50, Lr52, Lr60, Lr63, and
Lr67) as molecular markers controlling the leaf rust resistance in the selected Egyptian wheat varieties. The primer names, sequences, annealing temperature and size of the amplified fragment for
Lr genes markers used in this study are shown in
Table 3.
The molecular characterization results revealed that the 28
Lr tested genes yielded a total number of 34 scorable bands/amplicons. Out of the 28
Lr genes, nine
Lr genes were found to have more than one allele (
Lr13, Lr20, Lr24, Lr27, Lr37, Lr39, Lr47, Lr63, and
Lr67). Interestingly, six
Lr resistance genes (
Lr22a, Lr28, Lr29, Lr32, Lr34, and
Lr47-1) were found to be represented in all the studied varieties or, in other words, revealed a monomorphic pattern between all varieties. Notably, the
Lr10 resistance gene was considered the only unique positive marker gene since it showed a scorable band only in the BanySwif 5 variety, while it was absent in all of the other 49 studied varieties (
Table 4).
In terms of polymorphism levels, all primers showed a high percentage of polymorphism, except
Lr22a,
Lr28,
Lr29,
Lr32, and
Lr34, which showed absolute monomorphism. A narrow range of the expected heterozygosity values (H) were observed; the values were ranged between 0.0 to 0.5. A total of 9 out of the 28 primers showed very close and similar values near to 0.5. Moreover, the polymorphism information content (PIC) also exhibited the same range with values ranged from 0.0 to 0.5. The effective multiplex ratio values were more varied comparing to H and PIC; their values ranged from 0.02 to 1.94. On the other hand, the marker index and mean heterozygosity values were very low (ranging from 0.0000 to 0.00569). The discriminating power values ranged from 0.0 to 1.0. Additionally, the resolving power values were ranged between 0.0 and 2.12. The primer
Lr27 showed the best resolving power among all the used
Lr primers (
Table 5).
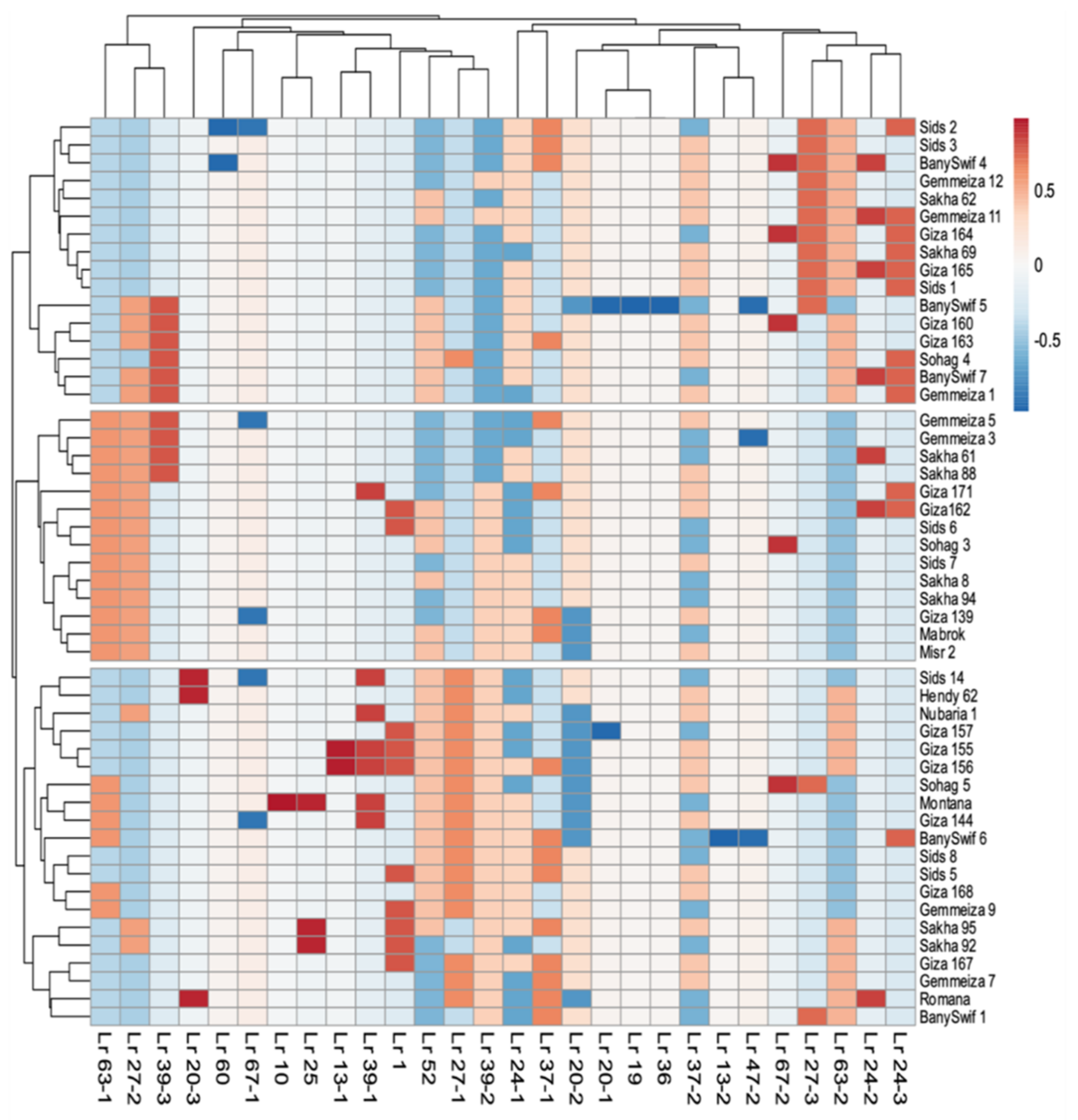
On the other side, the two-dimensional heatmap visualization of the interaction between the presence of
Lr genes and varieties performance revealed grouping of the 50 wheat varieties into three distinct groups (
Figure 1).
For principal component analysis (PCA), a scatter plotting of principal component 1 (PC1) plotted against principal component 2 (PC2) successfully separated the 50 wheat varieties into three sharp groups: (1) complete resistance varieties (red; Mabrok, Sakha 95, Montana, Sohag 5, Misr 2 and Giza 168), (2) fast rusting varieties (blue; Gemmeiza 1, 9, Giza162, 163, 164, 165, Sids 1, 2, 3, Sakha 62, 69, Sohag 3 and BanySwif 4), and 3) slow rusting resistance (green; Nubaria 1, Giza 139, 144, 155, 156, 157, 160, 167, 171, Sakha 8, 61, 88, 92, 94, Sids 5, 6, 7, 8, 14, Gemmeiza 3, 5, 7, 11, 12, BanySwif 1, 5, 6, 7, Romana, Hendy 62 and Sohag 4) (
Figure 2).
Dendrograms based on UPGMA analysis of
L r markers data were constructed for the 50 wheat varieties (
Figure 3). The dendrogram comprised two main clusters. The first cluster comprised only the Sakha 62 variety, while the second cluster comprised the other 49 wheat varieties. Particularly, the second cluster was subdivided into two main sub-clusters; the first sub-cluster comprised only the Sakha 69 variety, while the second sub-cluster was subdivided into three separate groups (
Figure 3).
3. Discussion
Rusts are one of the most destructive biotic stress in wheat and represent significant production constraints to wheat crop productivity worldwide. It has caused significant yield losses (about 60%) and diminished the quality of wheat grains. Three distinct types of rust diseases attack wheat: leaf rust (LR), yellow rust (YR), and stem rust (SR). These rusts are caused by certain pathogen species, consequently having many pathotypes that parasitize certain wheat varieties. Leaf rust disease caused by the fungus
Puccinia triticina (Eriks) is the most common bread wheat rust, causing significantly massive yield losses in wheat crops worldwide [
57]. One of the essential steps in which molecular marker techniques are used in developing efficient wheat breeding programs for rust resistance is determining and characterizing the wheat genotypes for their carrying of leaf rust resistance genes.
During the last decade, the dramatic development of molecular marker techniques and gene identification has facilitated the establishment of effective marker-assisted selection (MAS) systems, particularly towards wheat breeding for leaf rust resistance. These MAS systems were successfully established due to PCR-based markers’ availability for almost 80 designated leaf resistance genes/alleles [
23]. Remarkably, few studies have been released during the last decades describing the wheat germplasm that carries
Lr genes, especially in Egypt. This apparent lack of knowledge might be attributed to the giant genome of wheat and the existence of various pathotypes (races) that attack only certain varieties of wheat [
58].
Therefore, this study aimed to evaluate 50 Egyptian wheat varieties’ performance under open field conditions against leaf rust disease for two growing seasons (2018/2019 and 2019/2020) at the adult plant stage. The area under the disease progress curve (AUDPC) was used as the most reliable and convenient estimator to accurately measure the amount of rust infection. Based on the AUDPC, the evaluation results of these 50 varieties indicated that most of these varieties exhibited high to moderate leaf rust resistance, except varieties Gemmeiza 1, Gemmeiza 9, Giza162, Giza 163, Giza 164, Giza 165, Sids 1, Sids 2, Sids 3, Sakha 62, Sakha 69, Sohag 3, and BanySwif 4, which exhibited highly susceptible or fast leaf rusting behavior. Similar results were reported by Fahmi et al. (2015) [
59], and Pathan and Park (2006) [
60], who found that the Giza 163, Giza 164, Sids 1, and Sakha 69 wheat varieties were highly susceptible, compared to slow-rusting cultivars. The durability of resistance is supported by the diversity of resistant genes.
On the other side, we characterized these 50 varieties for their leaf rust resistance genes (28
Lr genes). Out of these 28
Lr genes, 21
Lr genes (
Lr1,
Lr10,
Lr13,
Lr19,
Lr20,
Lr22a,
Lr24,
Lr25,
Lr27,
Lr28,
Lr29,
Lr32,
Lr34,
Lr36,
Lr37,
Lr39,
Lr47,
Lr52,
Lr60,
Lr63,
Lr67) were successfully identified. Out of 15
Lr genes reported conferring the adult plant resistant or slow rusting behavior in wheat [
16,
61,
62], only five genes (
Lr13,
Lr22a,
Lr34,
Lr37 and
Lr67) were observed within the 50 Egyptian wheat varieties. The genes
Lr13,
Lr19,
Lr20,
Lr22a,
Lr28,
Lr29,
Lr32,
Lr34,
Lr36,
Lr47, and
Lr60 were the most predominant leaf rust resistance genes recognized across the 50 Egyptian wheat varieties. Among these genes, the
Lr13 gene was previously reported as the most broadly distributed
Lr gene worldwide [
63]. Moreover, it has been described that across European wheat genotypes, 58% of these tested genotypes were found to carry the
Lr13 gene alone or in combination with other resistance genes [
60,
64]. Moreover, Australian wheat genotypes were also found to contain this gene singly or combined with other race-specific genes [
65].
Regarding
Lr34, as expected, we identified this gene in almost all test Egyptian varieties, which is in good accordance with previous reports of Singh and Rajaram (1992) [
66], Imbaby et al., (2014) [
67] and Fahmi et al. (2015) [
59]. For
Lr37, which has been reported to mainly confer the adult plant’s resistance rather than in seedlings, it was found to be represented in about 60% of the tested Egyptian wheat genotypes. These results agreed with previous reports of Imbaby et al. (2014) [
47] on Egyptian germplasm and Singh and Rajaram (2002) [
65] on Western European germplasm. Meanwhile,
Lr19 and
Lr24, which were reported to be genetically linked to stem rust resistance genes
Sr25 and
Sr24, were found to be represented in about 98% and 66% of the tested wheat genotypes, respectively.
For
Lr28 and
Lr29, it was previously observed in 5 and 10 Egyptian wheat varieties, respectively [
5]. They also found that these varieties carried the
Lr25 and
Lr67 genes that might explain their higher degree of resistance. This finding was in complete agreement with our obtained results which confer that the
Lr28 and
Lr29 genes were represented in all the tested Egyptian wheat genotypes.
From another perspective, the molecular phylogeny analysis of the Egyptian wheat collection revealed accordance classification with the field evaluation of leaf rust resistance results with a little dissimilarity. The dendrogram gathered the genotypes characterized by high to moderate leaf rust resistance in one cluster while keeping those who exhibited highly susceptible or fast leaf rusting performance in a separate cluster, with a few exceptions.
The marker-assisted selection approaches grant the opportunity to select lines with desirable traits based on their genetic constituents rather than phenotypic performance, especially those combining several genes in a single genotype. With the guidance of molecular marker techniques, the pyramiding breeding of
Lr genes (even those functional at the seedling and/or adult plant stages) is expected to facilitate the designing of efficient breeding programs for durable resistance against this pandemic wheat disease. Undoubtedly, the resistance mechanisms against leaf rust are still poorly understood, but the information gained from resistance genes that are found in many varieties can help breeders to develop resistant varieties [
67]. This could be the most ecological and economical solution to manage wheat rust disease [
8]; additionally, the resistance durability seems to be dramatically improved when Lr resistance genes are combined [
68], whereby when more LR genes are accumulated in a variety, the combined effects of these genes give this variety a large base to resistance to disease [
8].
Therefore, molecular markers can be used effectively to confirm the existence of desired Lr resistance genes within the genetic background of certain wheat varieties and, consequently, to choose the most appropriate parents for efficient breeding programs.
5. Conclusions
The mining, characterization, and distribution of Lr genes within certain wheat genotypes/collections are crucial for developing new wheat-resistant genotypes. Gene pyramiding of Lr genes with the aid of molecular markers is necessary for ensuring the long-term sustainability of leaf rust resistance in Egyptian wheat varieties. In this study, 50 Egyptian wheat varieties were evaluated for their leaf rust resistance level at the adult plant stage for two successive seasons. The evaluation results indicated that most of the Egyptian wheat collection (37 out of 50 varieties) demonstrated high to moderate leaf rust resistance levels. Additionally, out of 28 Lr genes screened within the wheat collection, 21 Lr genes were successfully observed. Distinctly, 11 Lr genes (Lr13, Lr19, Lr20, Lr22a, Lr28, Lr29, Lr32, Lr34, Lr36, Lr47, and Lr60) were characterized as the most predominant Lr genes within the 50 Egyptian wheat varieties. Ultimately, our findings can act as a fundamental base for successful and efficient breeding programs for leaf rust resistance in an Egyptian wheat collection.


 ,
, 




{kind=link}
{kind=link}
{kind=link}