1. Introduction
Since plant breeding’s inception, the primary goal has been to increase the agronomic yield of cereal crops utilized as major food sources. This is readily apparent in the history of common wheat (
Triticum aestivum L.) improvement where the total production has increased globally by 0.9% from 1961 to 2008 [
1]. These yield improvements primarily represent the genetic gains achieved via selection for adaptation to local environments but also include an increased production area. Continuing to increase the common wheat production area will become more difficult due to a scarcity of land suited to agriculture and changes in the rainfall patterns associated with climate change [
2]. The percent of global agriculture acreage devoted to common wheat production has been as much as 21% [
3]. In the 2020/2021 growing cycle, global wheat production totaled 776 million metric tons, produced across 223 million hectares (USDA 2022). The world’s population is projected to continue increasing, such that by 2050 it will be necessary to produce roughly twice as much food to sustain the global population as compared to 2010. This will require an improvement in yield averaging 2.4% per year [
1], far above the current 0.9% increase per year for wheat. It is thus essential to focus on the identification of genetic variants that can be incorporated into new varieties that can increase wheat yield, since doubling the amount of arable acreage may not be possible.
When seeking to enhance the common wheat yield, plant breeders have been primarily focused on improvements to individual grain weight (GW) and grain number (GN). GW refers to the average weight of an individual grain and often appears in agronomic studies under the metric thousand kernel weight (TKW). GN represents the number of grains produced per wheat head. Agronomic yield is, therefore, a function of GW times GN times the number of productive heads over a specific area. There are numerous genes known to directly impact the number of productive heads within a specific unit area, such as
Teosinte branched 1 (
TB1), which exhibits an effect on tillering, but they are not the focus of this review [
4]. The focus here is on the genetic control of both the GW and GN. It is well documented that these two traits are negatively correlated, where increases in GN are associated with decreases in GW [
5,
6,
7]; however, many studies have shown a transgressive segregation for yield when crossing high GN and high GW cultivars [
8,
9,
10]. This trade-off is likely due to competition between grains as a sink, for a limited amount of source carbohydrates to “feed” the grain during grain fill [
11]. The observation of transgressive segregation for yield when crossing high GN and high GW cultivars indicates there is room to improve the yield regardless of the negative relationship between these two traits. The trade-off between the GN and GW is an example of the dichotomy of the reproductive strategy and parental investment that exists in nature. Larger but fewer seeds represent more investment of resources for fewer individuals, and more numerous but smaller seeds represent less investment of resources into more individuals [
5]. Studies focused on GN and GW across different cereals and environments have determined that GN is a highly plastic trait, while GW is significantly more heritable [
5].
The following review will characterize some of the known genes that effect GW and GN, but it is necessary to point out that GW, GN, and yield are all polygenic traits. Many of the genes discussed in this review were shown to influence either the GN or GW through recombinant inbred line (RIL) experiments, only to find that they had no effect in subsequent experiments in different environments. Recent studies have demonstrated this genotype by environment interaction by repeating RIL experiments on a variety of known genes and quantitative trait loci (QTL) that effect the GN or GW, only to find significant results for less than half of the previously identified genes or QTL [
12,
13,
14]. This does not indicate that the genes discussed are not useful to increase yield, but rather that the magnitude of the effect will be different under different environments and in different genetic backgrounds. A recent analysis of 390 diverse cultivars revealed that a significant majority of the yield increase in modern wheat is due to increasing both the GN and the number of wheat heads per area, with little gains in GW [
15]. Given that most of the past improvement has been the result of increasing GNs, there is still a large amount of potential yield increase via increasing the GW. An analysis of 27 elite cultivars, grown in Valdivia, Chile, Ciudad Obregon, Mexico, and Colorado, USA showed a much smaller association between yield and GN in the USA as compared to Mexico and Chile, indicating that increasing the GW may be a more stable goal for yield improvement [
16]. There are further implications here with respect to genotype by environment interactions, with environments that favor wheat (warmer, wetter, and longer growing seasons) garnering better yields through a focus on increasing the GN, while less favorable environments (cooler, drier, shorter growing seasons) will see more stable yield improvements through a focus on increasing GW.
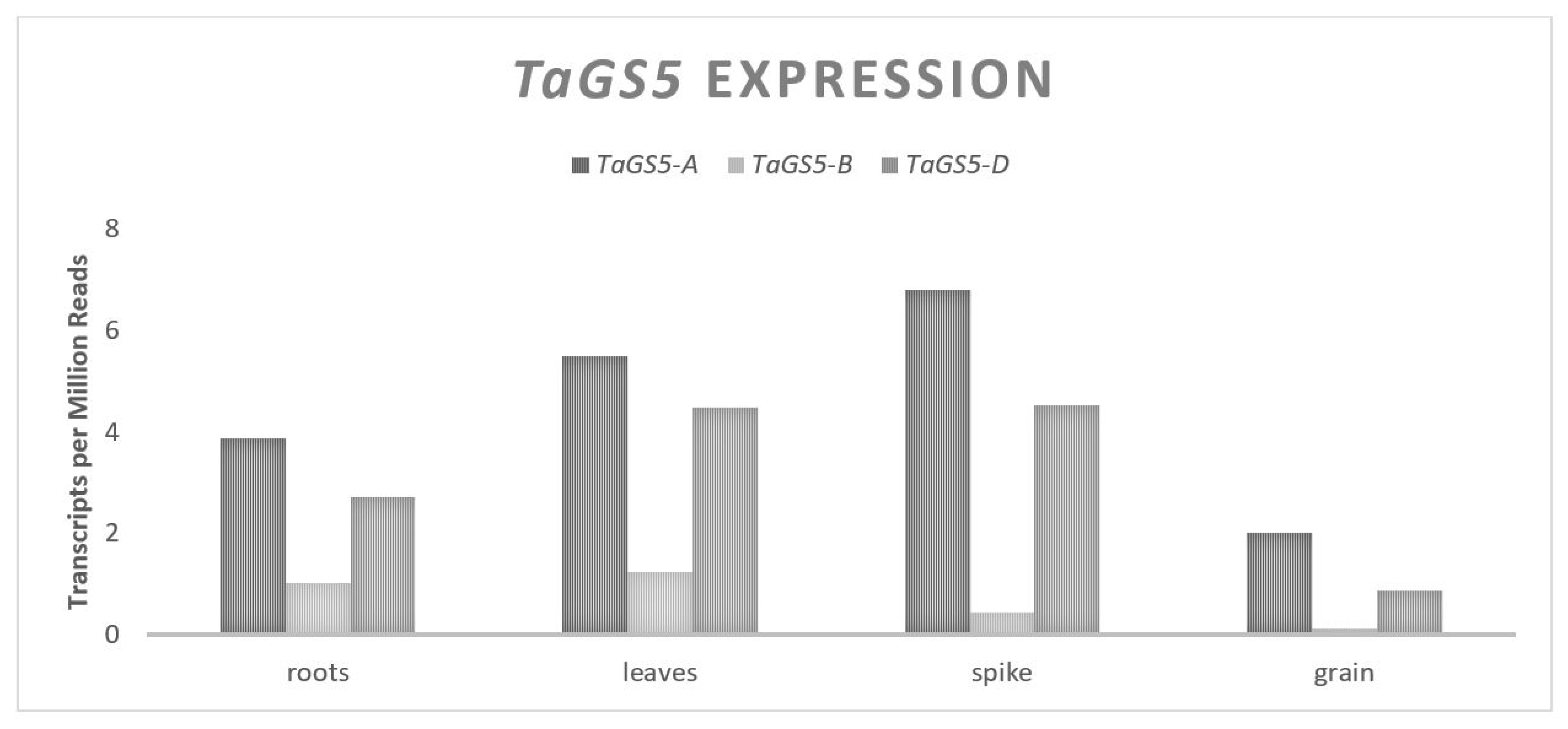
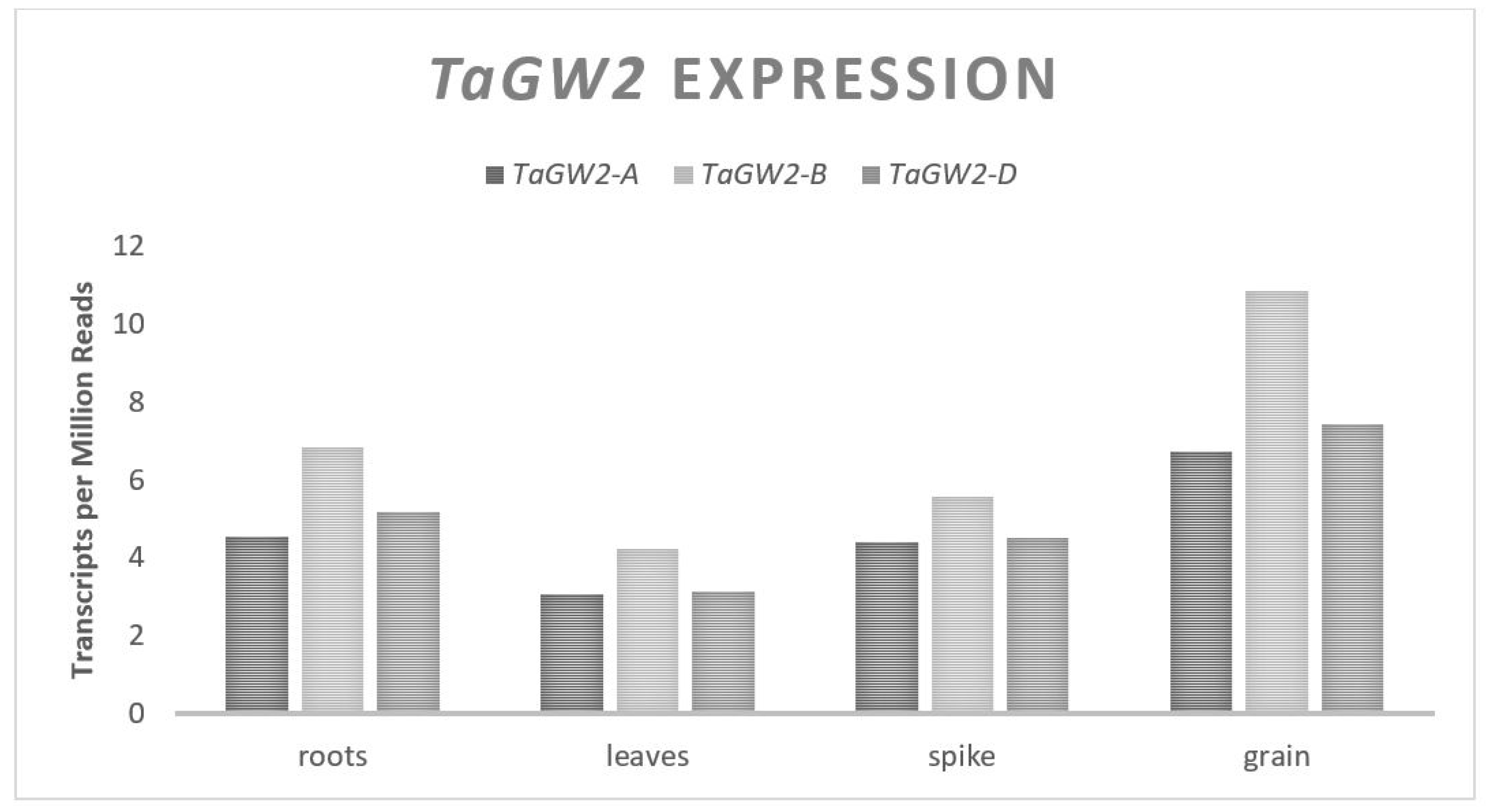
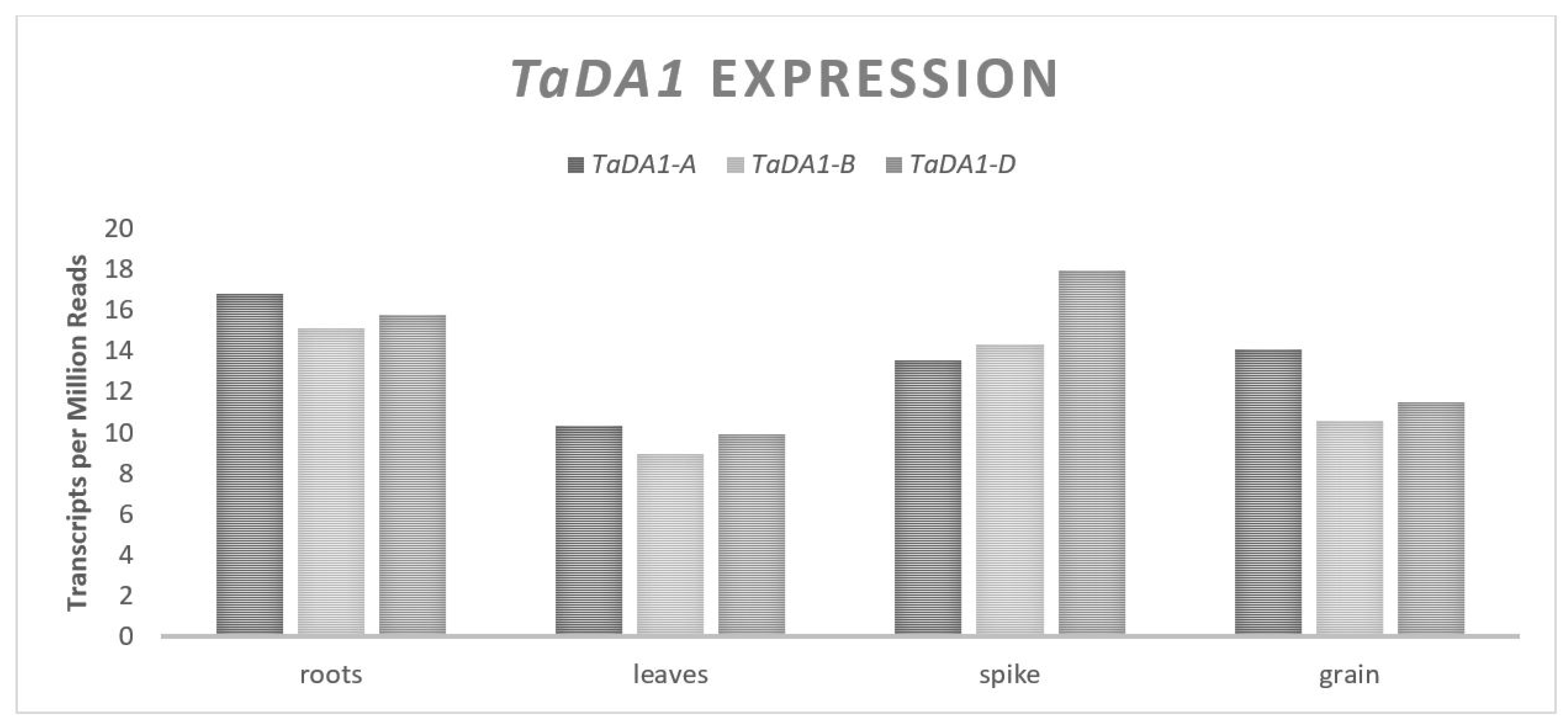
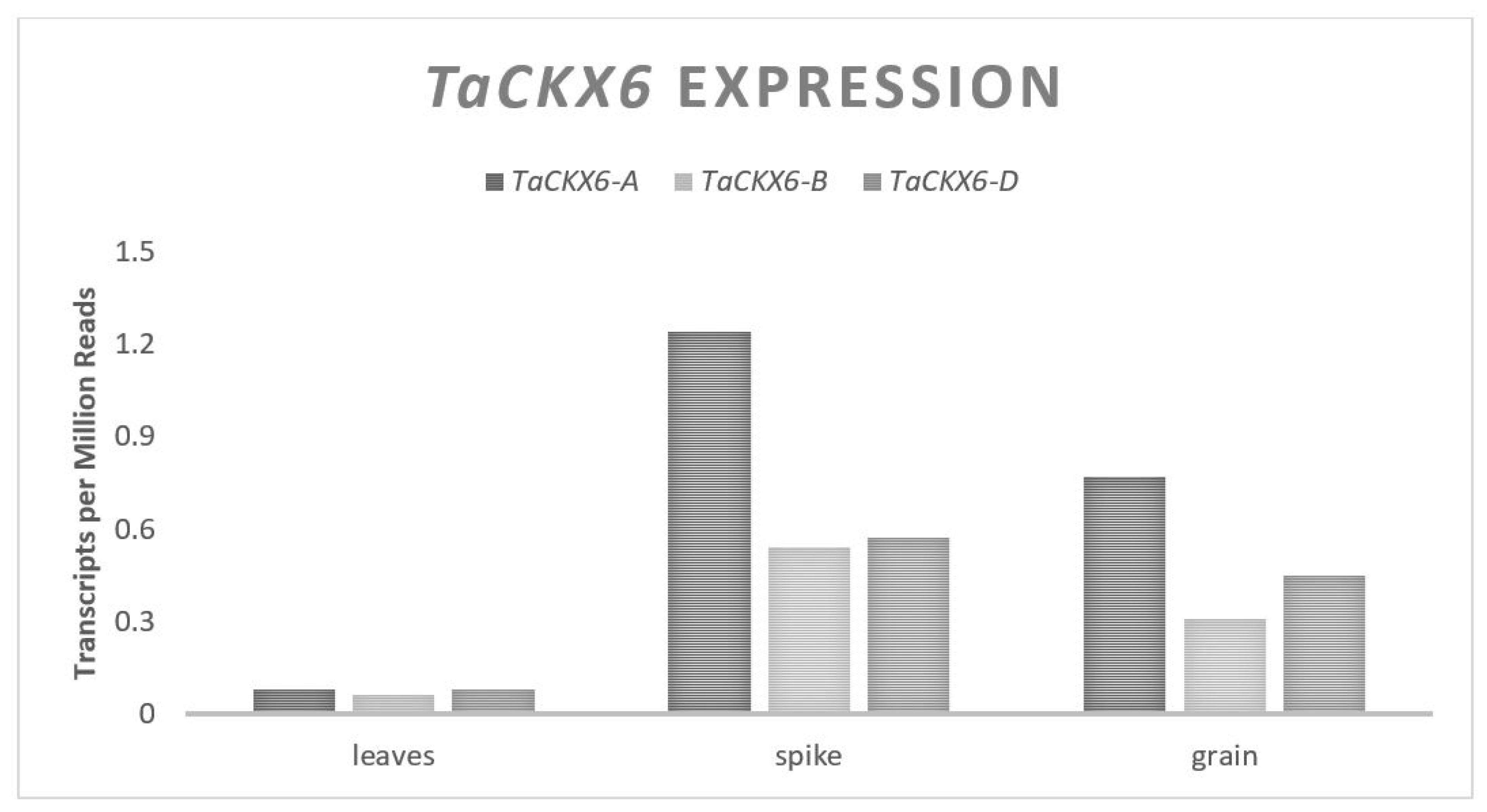
Three major mechanisms of control over the GW and GN will be discussed in this review, the first of which will focus on how plant size and the available carbohydrates affect the traits, addressing the source end of the previously mentioned trade-off. This is followed by an exploration of the genes that influence the size and number of pericarp tissue cells at the time of anthesis and their relationship to GW. Finally, genes that influence the floral architecture, floret fertility and their relationship to GN will be discussed. Given that most past yield improvements were the result of increases in GN per unit area, the genes that primarily affect GW will be the focus. Genes impacting the GW include a serine carboxypeptidase,
TaGS5, an E-3 ligase,
TaGW2, a ubiquitin receptor,
TaDA1, a cytokinin oxidase/dehydrogenase,
TaCKX6, and an expansin,
TaExp6 (
Table 1). An F-Box gene,
WAPO1, and a transcription factor,
TaGNI, that both affect the GN will be discussed as well (
Table 1). Primers necessary to sequence the genes in this review are included in
Supplementary Table S1. These genes were chosen because they each impact different plant development functions, alterations of which can be seen as different strategies for yield improvement. This review seeks to highlight how each of these strategies affects the source or sink tissue and makes recommendations for how agronomically preferable alleles of these genes might work best together to improve yield.
2. Plant Mass/Available Carbohydrates
The process of grain filling is informed by the well-established source-to-sink pathway that exists in plants. Plants utilize photosynthesis to build sugars which are in turn mobilized to the tissues in the plant for use in either new growth or are stored in seeds. The photosynthetic tissues represent the source while the grains represent the sink. In order to determine whether wheat GW was source or sink limited, researchers removed spikelets from different positions in wheat heads at anthesis and observed how the GW was impacted. No significant difference was observed in GW between the plants with the spikelets removed and the plants left fully intact [
49]. The consistent GW measured between the plants with and without the spikelets removed suggests that there is not sufficient competition between grains for the available assimilates to limit their potential size. The implication is consistent with the fact that most yield enhancements over the past century have come by increasing the GN and reinforce the idea that GW is more sink than source limited. Subsequent repeat experiments across multiple environments showed the same results, indicating that the environment does not have a strong impact on wheat grain fill and that GW is more sink than source limited [
50].
Regardless of the likelihood that grain fill is more limited by sink than source, there is evidence that increasing the source is associated with a higher GW. The source potential is largely made up of two different plant properties. First is the area of photosynthetically active tissue which directly impacts the amount of sugars the plant can produce. Second is the concentration of water soluble carbohydrates (WSC) that can be mobilized during grain fill as the photosynthetic tissues of the plant senesce [
51]. While the environment can impact the WSC, an analysis of 116 diverse lines in four different environments revealed that differences in the stem WSC content were largely (90%) explained by genotype [
52]. Researchers compared the GW of a RIL population, derived from the cross of a high and low WSC cultivar, finding a positive association between high WSC and GW [
53]. The increased availability of WSC being associated with a higher GW contradicts the idea that source is not limiting for wheat during grain fill; however, the same study found that high WSC lines were associated with a smaller GN. There is a strong positive association between the photosynthetic material above the flag-leaf node and the GW in wheat [
54], the implication being that enhanced source material leads to an enhanced GW. This suggests again that there may be some source limiting properties to grain fill. Early studies on the
Reduced Height 1 gene
Rht-1, which is most well-known for controlling plant height, found a direct association between taller plants and a higher GW [
55]. As the plants were reduced in height by mutations in different
Rht homologues, so was their GW, indicating again that grain fill may be source limited. It is important to remember that while an increased plant size is associated with an increased GW, it is also associated with increased dry matter [
55]. The increased dry matter, composed of longer stems, roots and more tillers and leaves, can be considered ancillary material that may be a competitive sink for grain development, especially under resource limited dryland conditions. Modern wheat breeding has frequently selected for mutant
Rht genes to reduce plant height, which results in reduced individual GWs but enhanced yields [
56]. This illustrates that while increased plant mass contributes to increased GW as well as ancillary tissues, the yields may also decrease.
Literature addressing the degree to which grain fill in wheat is source limited is contradictory at best. What can be definitively stated is that enhancing the source material, be it an increased photosynthetic flag-leaf area or higher WSC in the stem, is associated with a higher GW. Interestingly, there is an association between higher WSC lines and drought tolerance. It is theorized that under drought conditions grain fill is more dependent on the amount of WSC built up in the stem tissues made available during grain fill, as water limitations reduce the photosynthetic activity [
57,
58]. Conversely, high WSC are associated with an increased GW under irrigated conditions, likely due to the increased source for filling more numerous fertile florets tissues [
53]. These findings have implications for breeding, indicating that enhancing the source materials used in grain fill may be a good strategy for seeking yield improvements in irrigated environments. Additionally, breeding for dryland environments will be favored by genes that increase WSC over time, supporting the grain fill under conditions of limited photosynthetic activity.
5. Discussion
The first and foremost goal of breeding for globally utilized cereal grains has been and remains increasing the yield. Historically, efforts to improve wheat have focused on improving the yield, which has been predominantly accomplished by increasing the GN while keeping the GW constant, or vice versa [
15]. Frequently, breeding efforts to increase one trait result in a decrease of the other, while the yield itself is held constant [
5,
6,
7]. This is largely due to the phenomenon of the source-to-sink pathway that exists in all plants, in which carbohydrates are generated by source materials such as photosynthetic tissues, and mobilized to their final sink tissue, namely, the developing grain [
11,
49]. It therefore makes sense that the GN and GW are negatively linked as more grains represent more sinks to fill, frequently resulting in fewer assimilates making it into individual grains. This relationship is not one to one, and crosses between high GN and high GW lines often display transgressive segregation for the respective traits, as well as yield [
8,
9,
10]. The continued reporting of transgressive segregation for such crosses indicates that GN and GW can still be improved independent of each other.
Improvements to the source side of the source-to-sink pathway appear to have a direct positive effect on GW, generally independent of the GN. By enhancing the source, the potential for more photo assimilates to be mobilized to developing grains becomes apparent. Enhancements to the source potential should have little effect on the GN, at least at the level of genetic control [
53,
54,
55]; however, increased availability of the resources within the plant may ultimately lead to fewer floret abortions, allowing for an increase in the GN along with GW [
53]. Several strategies have been highlighted for improving the source potential of the wheat plant, first and foremost being the plant size. Larger plants are associated with larger grains and plant size can be affected in a large magnitude by a small number of genes, such as
Rht [
55]; however, controlling the plant height is important in wheat breeding for different reasons other than control over the GW, such that it is not always advantageous to breed for the largest plants. The established positive relationship between the flag-leaf photosynthetic area and GW indicates that exploiting the genetic control over the flag-leaf size is a reasonable strategy for increasing the sink potential, and enhancing the GW without decreasing the GN [
54]. Similarly, the association between higher stem WSC and a higher GW is apparent, such that the selection for lines with increased WSC represents an improvement to the plant source potential [
53]. Utilizing awned genotypes as opposed to those without represents an increased investment into the source potential of the plant, as the awn photosynthetic tissue directly moves assimilates into the developing grains; however, the development of awns likely represents a competitive sink with developing maternal tissues and is associated with a decrease in the GN as a result [
79,
80,
81,
82]. While awns represent an increase to the source potential, they may simultaneously decrease the sink potential and are, therefore, a less useful trait in breeding for a stable yield increase. Selecting for awned versus awnless lines does have utility depending on the target environment.
Given that much of the gain in yield since the Green Revolution has come from increasing the GN per unit area [
15], stable GW improvement is potentially a more fruitful direction to find the necessary yield improvements required to achieve the goal of doubled production by 2050 [
1]. A major area of genetic control over GW comes in promoting more numerous and larger pericarp cells at the time of anthesis [
59,
60]. This can be thought of as improvements to the sink potential of the plant and there are many genetic pathways in the plant that can be manipulated to enhance the pericarp cell proliferation. Control over the G1/S phase genes can be exploited to promote pericarp cell proliferation [
17,
18,
19,
61]. Increasing both the cytokinin and expansin expression in the developing wheat head, leading up to the time of anthesis, also promotes larger and more numerous pericarp cells [
37,
40,
41,
42,
62,
63,
69]. Allelic differences in the genes that control the synthesis and degradation of cytokinin and expansin in the wheat head leading up to anthesis can be exploited to promote pericarp cell proliferation. Increasing the cytokinin and expansin levels in the plant leading up to anthesis does more than just improve the sink potential of pericarp tissues, it also enhances the source potential of photosynthetic tissues given the association between increased cytokinin levels in the cell and increased chlorophyl production [
34,
63]. This may explain why experiments that increase cytokinin or expansin levels in the cell frequently see stable GW improvements without GN decreases. A proportional increase to the source potential, alongside the increase to the sink potential may be essential in stable yield improvements outside of the normal GW/GN trade-off (
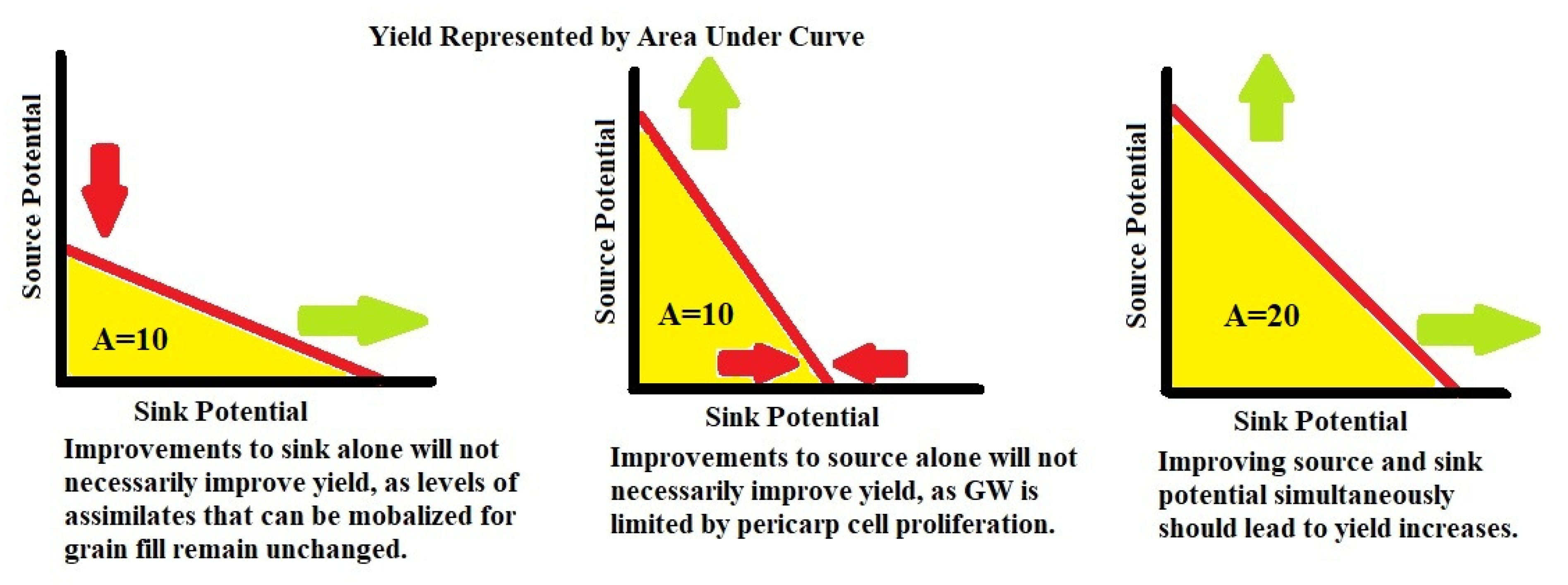
Figure 7).
Genes that control the floral architecture and floret fertility also contain the potential for a stable yield improvement.
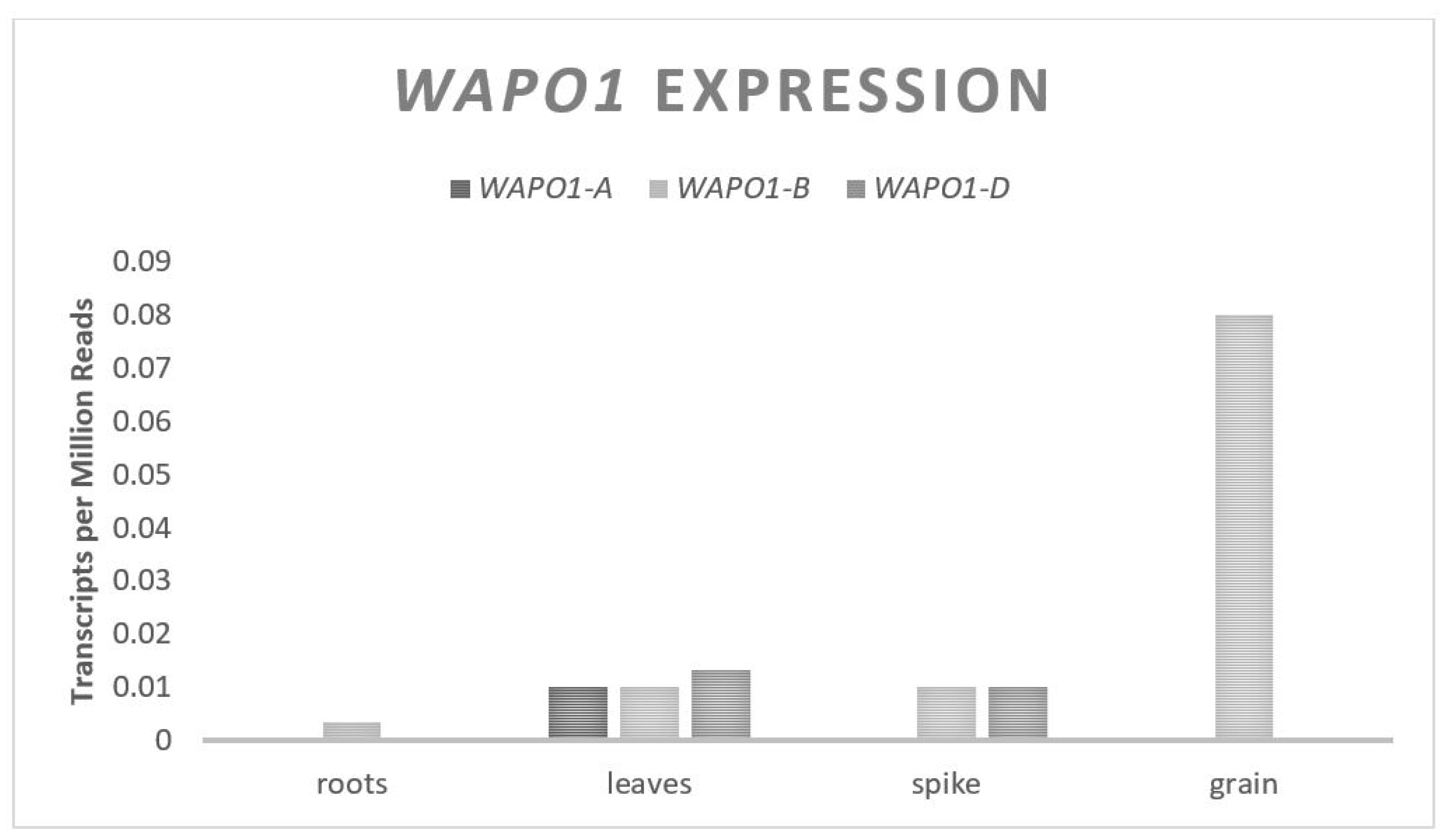
WAPO1 directly impacts the floral architecture with a higher expression leading to more spikelets and a higher GN; however, this is coupled with a higher rate of floret abortion [
44].
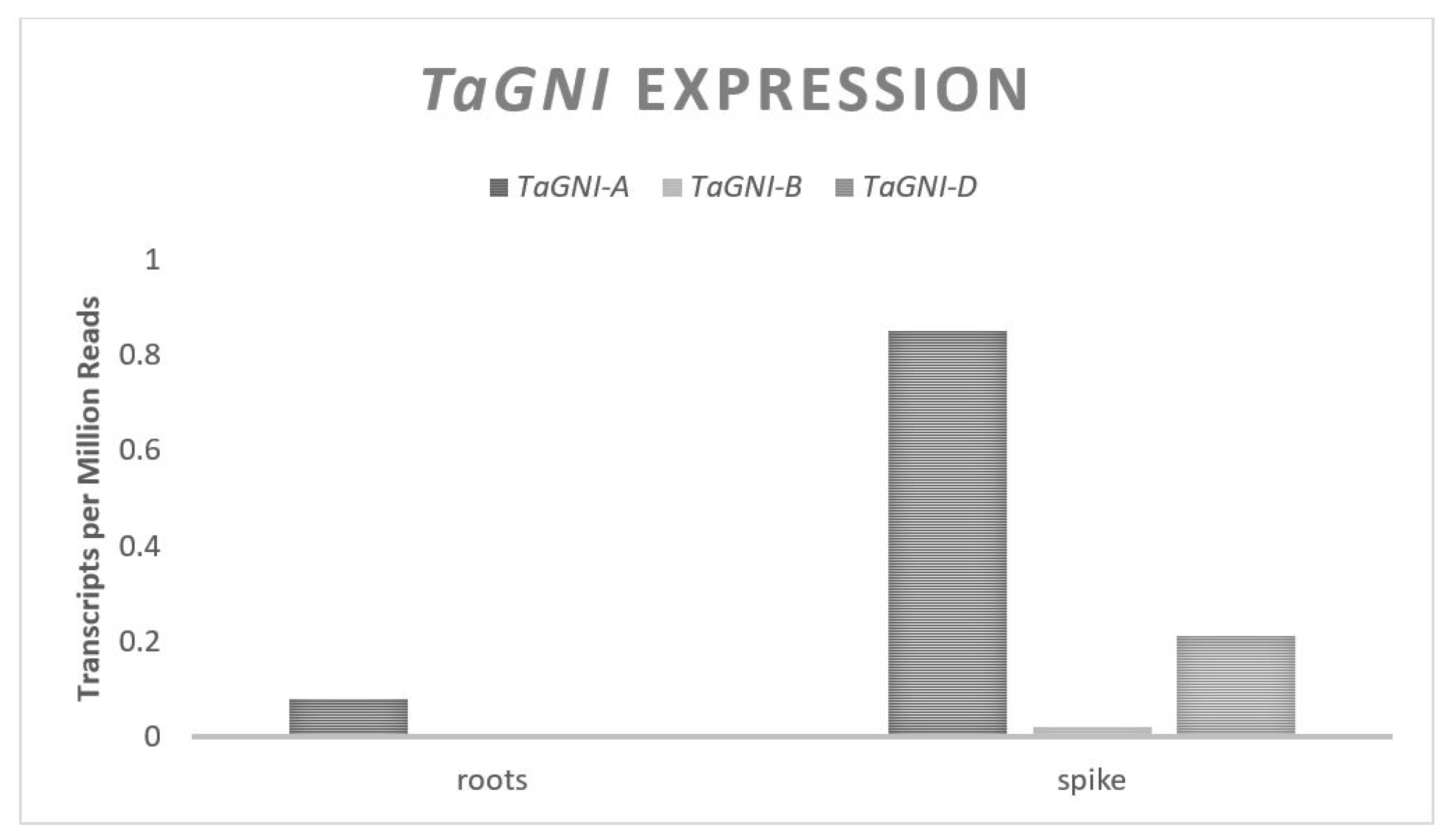
GNI directly impacts floret abortions regardless of the environment, with less functional alleles resulting in a larger GN [
47,
48]. Combining alleles of
WAPO1 that promote more spikelets with alleles of
GNI that reduce floret abortion may be useful in further enhancing GN. It is still possible for a larger number of grains to have a smaller GW as a result of the source-to-sink relationship, but this decrease can potentially be mitigated by additionally selecting for alleles of genes that directly enhance GW through the promotion of larger pericarp cells at anthesis (
Figure 8). The frequency with which crosses between a high GN and high GW cultivar adapted to an area display transgressive segregation [
8,
9,
10], is a good indicator of a strategy to identify the genes that may lead to the most stable yield gains. The recommendations for breeding would be to perform such a cross with locally adapted cultivars and utilize a Genome Wide Association analysis on the resulting population to identify which genes are associated with the most improvement to GN or GW in the adapted environment.
Genotype by environment interaction plays a large role in determining the best method by which to pursue a yield enhancement [
12,
13,
14]. There are a wide variety of growing environments around the world, but they can be aggregated into two distinctly different situations. There are irrigated environments, in which the nutrients and water necessary to grow and maintain the plant are always or more frequently available as opposed to dryland environments, where the resources are limited. Dryland or resource limited environments can, therefore, be thought of as limiting to the source potential of a plant, while irrigated environments are significantly less limited in this regard. Given this understanding, it would be more advantageous to focus on the development of source potential through genetic control when breeding for resource limited environments, and placing more focus on developing the sink potential through genetic control when breeding for irrigated environments. This idea is reinforced by the fact that high WSC lines are associated with more drought tolerance, as the increased abundance in WSC in the plant tissues can be mobilized for grain fill even during periods of reduced photosynthesis under water stress [
57,
58]. In resource limited environments it would be recommended to breed for lines with high WSC levels in the stem tissues that persist past anthesis into the grain fill. The source potential can be further increased by utilizing awned lines and selecting for a larger flag-leaf area [
79,
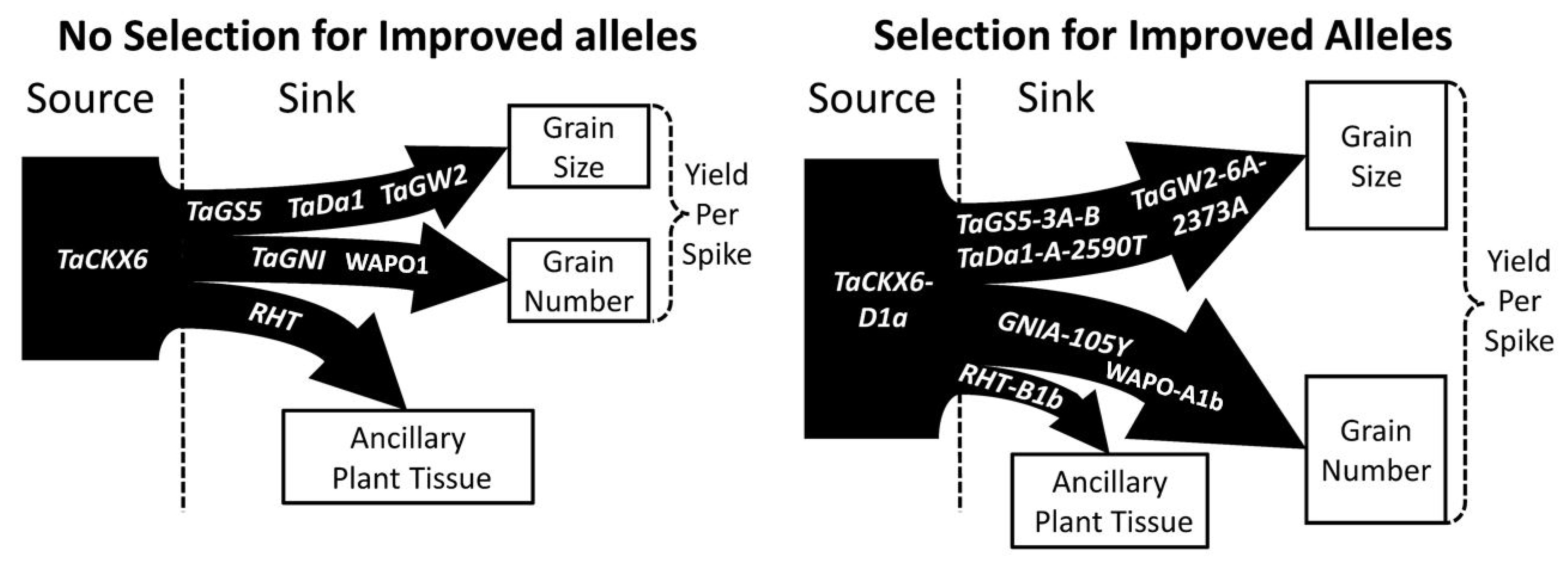
81]. Secondary to the efforts to improve the source potential of the plant at the genetic level, would be control of the size of the pericarp tissue through selection of the proper alleles of
TaGS5, TaGW2, TaDA1, TaExpA6, and
TaCKX genes (
Figure 9); however, in a source limited environment, adjusting the potential of the sink may not result in a stable yield increase [
16]. With that in mind, it would also seem appropriate to select for alleles of
WAPO1 that promote fewer spikelet formations, such as
WAPO-A1a, and alleles of
GNI that promote floret abortion, such as the wild type
GNI-105N allele, so that in a source limiting environment, fewer sinks are competing for assimilates during the grain fill (
Table 8).
Irrigated environments have significantly less source limitations, as the availability of water should drive photosynthesis consistently throughout the life cycle of the plant, provided there is sufficient fertilizer. While lines with increased WSC may result in a benefit to the yield in irrigated environments, the fact that these environments are significantly less source limiting decreases the potential impact of a high WSC line on the yield [
53], suggesting that other breeding strategies may be more effective. With less source limitations, breeding for a higher GN still represents a potential for increasing the yield, therefore, strategies should include the selection of alleles of
WAPO1 that promote spikelet proliferation, such as
WAPO-A1b, and alleles of
GNI that decrease the floret abortion, such as
GNI-105Y. Increases in GN should be coupled with the selection of alleles of
TaGS5, TaGW2, TaDA1, TaExp06, and
TaCKX genes that increase the GW [
18,
26,
29,
32,
38,
41] (
Table 9). In particular, the alleles of
TaGS5,
TaExp06, and
TaCKX genes that promote increased pericarp cell size and numbers would likely be most useful, as their mechanism for promoting a higher GW comes through the control of cellular division and elongation [
37,
41,
61].
TaGW2 and
TaDA1 also affect the cytokinin levels in cells, which in turn impact cellular division; however, they also impact the levels of AGPase expression [
26,
63]. Alleles of these two genes can be selected to promote a higher GW, but may increase the GW by mobilizing more starch to the grain [
65]. This could affect the end-use quality of the grain, specifically the protein content, in such a way as to make the final grain less economically useful. More studies on how the allelic variation of
TaGW2 and
TaDA1 effects the protein content of the final grain are warranted in order to determine the best strategy for exploiting these loci for germplasm improvement.
The trade-off between GN and GW is often thought of as a simple competition for resources between sinks [
5,
6,
7]. In many cases this is true, but this review has highlighted multiple strategies for making yield improvements through the control of one or both of these traits in spite of this trade-off. The most obvious strategy calls for proportionally improving the source potential of the plant alongside improvements to the sink potential of the grain. Crop scientists have been charged with making rapid yield improvements to meet the growing demand and with fixing the most beneficial alleles of
TaGS5,
TaGW2,
TaDA1,
TaExp06,
TaCKX6 and
GNI into more modern cultivars that will go a long way in supporting this important endeavor.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}