
A Crop Modelling Strategy to Improve Cacao Quality and Productivity

 ,
,
Abstract
:1. Introduction
Floral Phenology of Cacao
2. Materials and Methods
2.1. Test Site and Yield Production
2.2. Weather Conditions
2.3. Inputs and Data Acquisition
2.4. Thermal Time for Pod Harvest Date Identification
2.5. Model Calibration
2.6. Parameters
2.7. Evaluation of Model Performance
3. Results
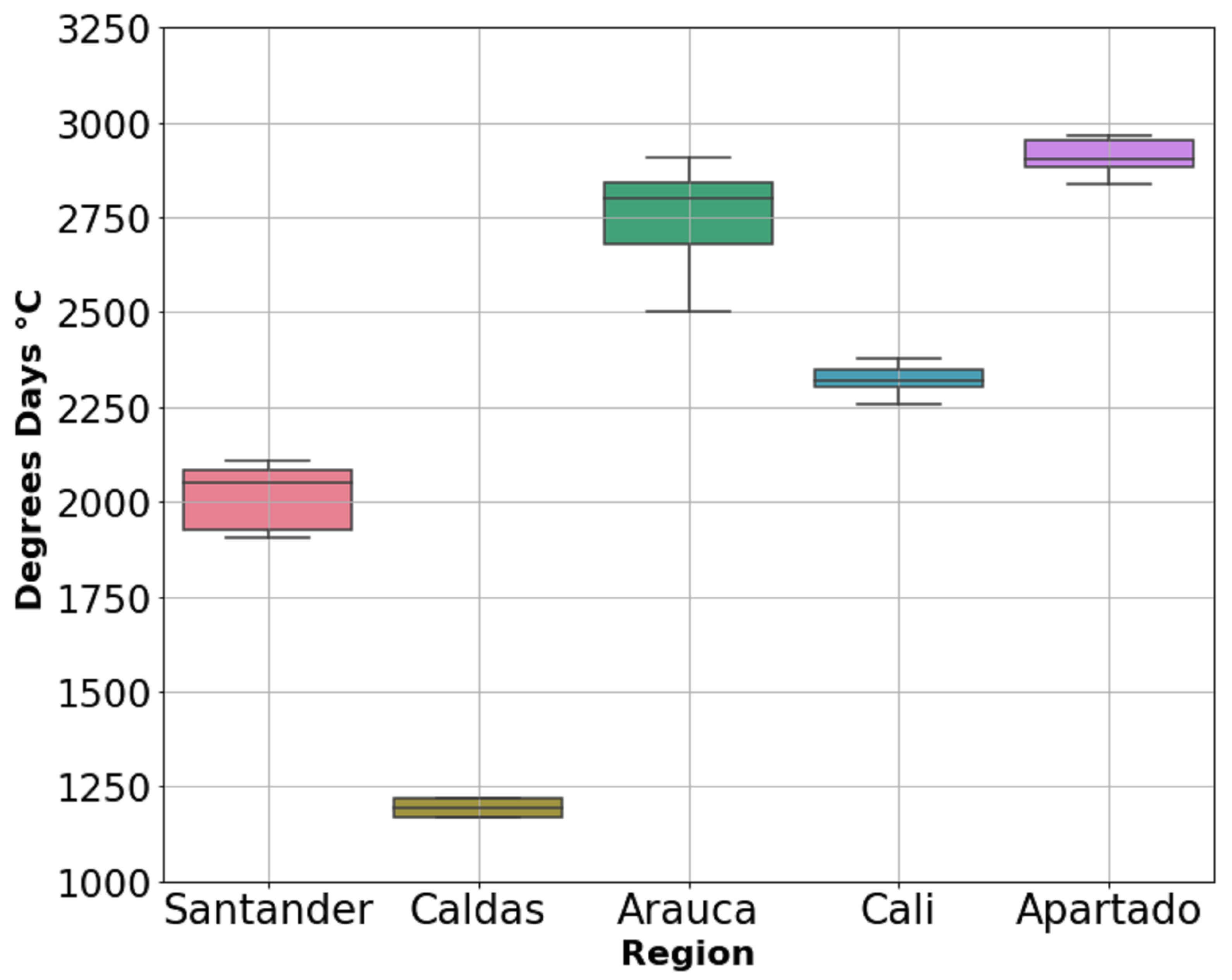
3.1. Weather Conditions over Flowering Time
3.2. Thermal Time
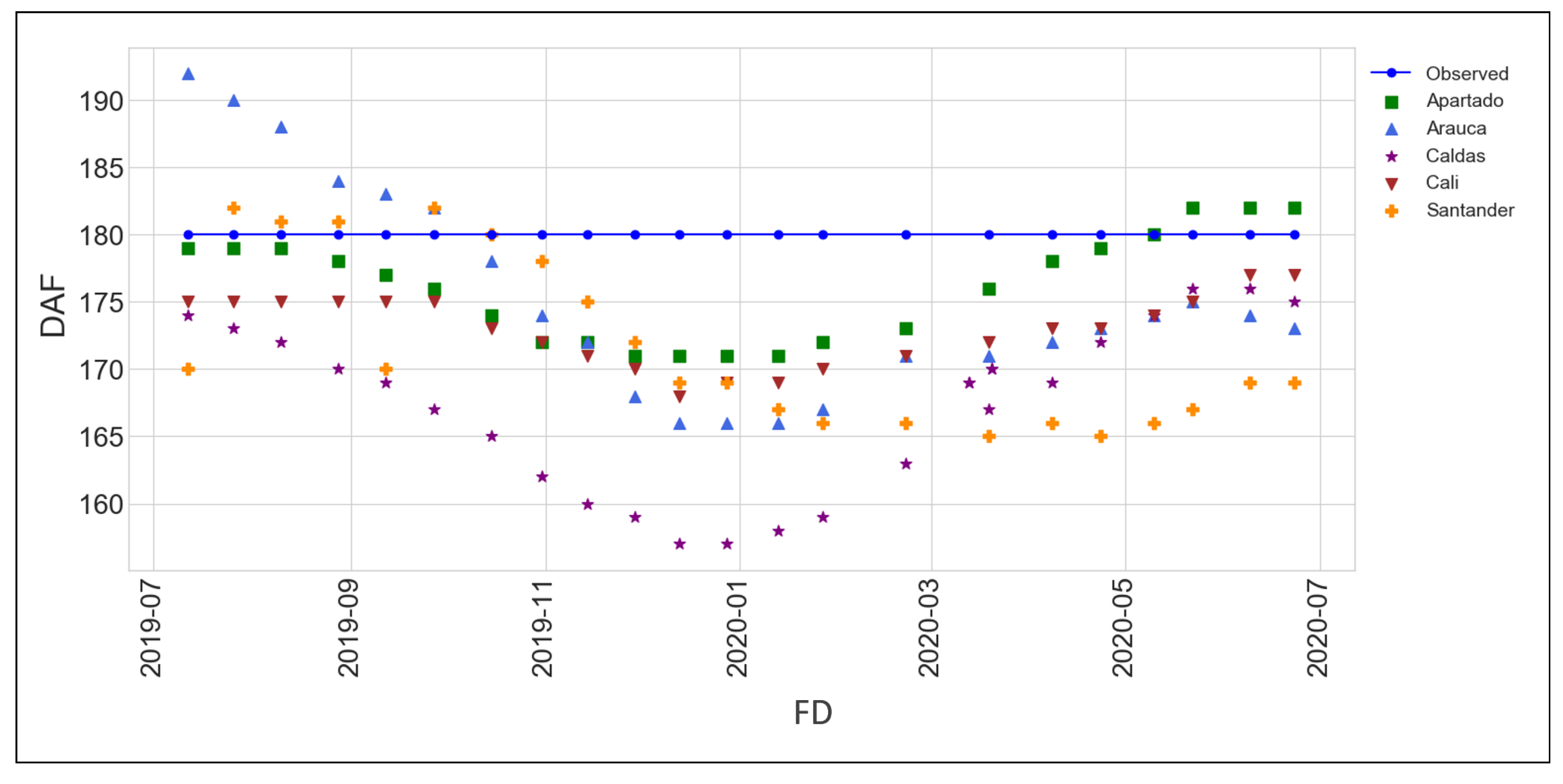
3.3. Model Validation
3.4. Predicting Optimal Pod Harvest Day
4. Discussion
4.1. Weather Effects over Flower Stability and Pollination
4.2. Thermal Time for Harvest Day Predictions
4.3. Cacao Crop Model Simulations
4.4. App Development
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuidema, P.A.; Leffelaar, P.A.; Gerritsma, W.; Mommer, L.; Anten, N.P. A physiological production model for cocoa (Theobroma cacao): Model presentation, validation and application. Agric. Syst. 2005, 84, 195–225. [Google Scholar] [CrossRef] [Green Version]
- Motamayor, J.C.; Risterucci, A.M.; Lopez, P.A.; Ortiz, C.F.; Moreno, A.; Lanaud, C. Cacao domestication I: The origin of the cacao cultivated by the Mayas. Heredity 2002, 89, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Argout, X.; Salse, J.; Aury, J.M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Medina, C.; Arana, A.C.; Sounigo, O.; Argout, X.; Alvarado, G.A.; Yockteng, R. Cacao breeding in Colombia, past, present and future. Breed. Sci. 2019, 69, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nix, A.; Paull, C.; Colgrave, M. Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review. Plants 2017, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Niemenak, N.; Cilas, C.; Rohsius, C.; Bleiholder, H.; Meier, U.; Lieberei, R. Phenological growth stages of cacao plants (Theobroma sp.): Codification and description according to the BBCH scale. Ann. Appl. Biol. 2010, 156, 13–24. [Google Scholar] [CrossRef]
- Suárez, Y.Y.J.; Castañeda, G.A.A.; Daza, E.Y.B.; Estrada, G.A.R.; Molina, J.R. Modelo Productivo para el Cultivo de Cacao (Theobroma cacao L.) en el Departamento de Santander; Corporación Colombiana de Investigación Agropecuaria—AGROSAVIA: Mosquera, Colombia, 2021. [Google Scholar] [CrossRef]
- Abbott, P.C.; Benjamin, T.J.; Burniske, G.R.; Croft, M.; Fenton, M.M.; Kelly, C.R.; Lundy, M.M.; Rodriguez-Camayo, F.R.; Wilcox, M.D. An Analysis of the Supply Chain of Cacao in Colombia; United States Agency for International Development: Washington, DC, USA, 2019. [Google Scholar]
- Gutiérrez García, G.A.; Gutiérrez-Montes, I.; Hernández Núñez, H.E.; Suárez Salazar, J.C.; Casanoves, F. Relevance of local knowledge in decision-making and rural innovation: A methodological proposal for leveraging participation of Colombian cocoa producers. J. Rural. Stud. 2020, 75, 119–124. [Google Scholar] [CrossRef]
- García-Cáceres, R.G.; Perdomo, A.; Ortiz, O.; Beltrán, P.; Lopéz, K. Characterization of the supply and value chains of Colombian cocoa. Dyna 2014, 81, 30–40. [Google Scholar] [CrossRef]
- Escobar, S.; Santander, M.; Useche, P.; Contreras, C.; Rodríguez, J. Aligning strategic objectives with research and development activities in a soft commodity sector: A technological plan for colombian cocoa producers. Agriculture 2020, 10, 141. [Google Scholar] [CrossRef]
- Lamos-Díaz, H.; Puentes-Garzón, D.E.; Zarate-Caicedo, D.A. Comparison Between Machine Learning Models for Yield Forecast in Cocoa Crops in Santander, Colombia. Rev. Fac. Ing. 2020, 29, 54. [Google Scholar] [CrossRef]
- Escobar, S.; Santander, M.; Zuluaga, M.; Chacón, I.; Rodríguez, J.; Vaillant, F. Fine cocoa beans production: Tracking aroma precursors through a comprehensive analysis of flavor attributes formation. Food Chem. 2021, 365, 130627. [Google Scholar] [CrossRef]
- Ceccarelli, V.; Fremout, T.; Zavaleta, D.; Lastra, S.; Imán Correa, S.; Arévalo-Gardini, E.; Rodriguez, C.A.; Cruz Hilacondo, W.; Thomas, E. Climate change impact on cultivated and wild cacao in Peru and the search of climate change-tolerant genotypes. Divers. Distrib. 2021, 27, 1462–1476. [Google Scholar] [CrossRef]
- Lahive, F.; Hadley, P.; Daymond, A.J. The physiological responses of cacao to the environment and the implications for climate change resilience. A review. Agron. Sustain. Dev. 2019, 39, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Medina, V.; Laliberte, B. A review of Research on the Effects of Drought and Temperature Stress and Increased CO2 on Theobroma cacao L., and the Role of Genetic Diversity to Address Climate Change; Bioversity International, CGIAR: Cartago, Costa Rica, 2017. [Google Scholar]
- Reynolds, M.; Kropff, M.; Crossa, J.; Koo, J.; Kruseman, G.; Molero Milan, A.; Rutkoski, J.; Schulthess, U.; Balwinder-Singh; Sonder, K.; et al. Role of modelling in international crop research: Overview and some case studies. Agronomy 2018, 8, 291. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, B.; Xiao, L.; Hoogenboom, G.; Boote, K.J.; Kassie, B.T.; Pavan, W.; Shelia, V.; Kim, K.S.; Hernandez-Ochoa, I.M.; et al. A SIMPLE crop model. Eur. J. Agron. 2019, 104, 97–106. [Google Scholar] [CrossRef]
- Bai, T.c.; Tao, W.; Zhang, N.n.; Chen, Y.q.; Mercatoris, B. Growth simulation and yield prediction for perennial jujube fruit tree by integrating age into the WOFOST model. J. Integr. Agric. 2020, 19, 721–734. [Google Scholar] [CrossRef]
- Romero-Vergel, A. Crop Modelling and Remote Sensing for Yield Predictions of Asparagus Cultivated in Peru; Ongoing Publication: Aberystwyth, Wales, 2022. [Google Scholar]
- López-Hernández, J.; López-Hernández, L.; Avendaño-Arrazate, C.; Aguirre-Medina, J.; Espinosa-Zagaroza, S.; Moreno-Martínez, J.; Mendoza-López, A.; Suárez-Venero, G. Biología floral de cacao (Theobroma cacao L.); criollo, trinitario y forastero en México. AGROProductividad 2018, 11, 129–136. [Google Scholar]
- Swanson, J.D. Flower development in Theobroma cacao L.: An Assessment of Morphological and Molecular Conservation of Floral Development between Arabidopsis thaliana and Theobroma cacao. Ph.D. Thesis, The Pennsylvania State University, State College, PA, USA, 2005. [Google Scholar]
- Cheesman, E.E. Fertilization and Embryogeny in Theobroma Cacao, L. Ann. Bot. 1927, 41, 107–126. [Google Scholar] [CrossRef]
- Berry, D.; Cilas, C. Étude génétique de la réaction à la pourriture brune des cabosses chez des cacaoyers (Theobroma cacao L) issus d’un plan de croisements diallèle. Agronomie 1994, 14, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Lehrian, D.W.; Keeney, P.G. Changes in lipid components of seeds during growth and ripening of cacao fruit. J. Am. Oil Chemists’ Soc. 1980, 57, 61–65. [Google Scholar] [CrossRef]
- Toledo-Hernández, M.; Tscharntke, T.; Tjoa, A.; Anshary, A.; Cyio, B.; Wanger, T.C. Landscape and farm-level management for conservation of potential pollinators in Indonesian cocoa agroforests. Biol. Conserv. 2021, 257, 109106. [Google Scholar] [CrossRef]
- NASA. Nasa Power. The National Aeronautics and Space Administration. 2021. Available online: https://power.larc.nasa.gov/ (accessed on 10 July 2021).
- RStudio Team. RStudio 1.4: Integrated Development Environment for R; RStudio, PBC.: Boston, MA, USA, 2020. [Google Scholar]
- Ritchie, J.T.; Nesmith, D.S. Temperature and Crop Development. In Modeling Plant and Soil Systems; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 1991; Chapter 2; pp. 5–29. [Google Scholar] [CrossRef]
- Slafer, G.A.; Rawson, H.M. Base and optimum temperatures vary with genotype and stage of development in wheat. Plant Cell Environ. 1995, 18, 671–679. [Google Scholar] [CrossRef]
- Daymond, A.; Hadley, P. Differential effects of temperature on fruit development and bean quality of contrasting genotypes of cacao (Theobroma cacao). Ann. Appl. Biol. 2008, 153, 175–185. [Google Scholar] [CrossRef]
- Erneholm, I. Cacao Production of South America. Historical Development and Present Geographical Distribution; CAB Direct: Gothenburg, Sweden, 1948; p. 279. [Google Scholar]
- Crout, N.; Craigon, J.; Cox, G.; Jao, Y.; Tarsitano, D.; Wood, A.; Semenov, M. An objective approach to model reduction: Application to the Sirius wheat model. Agric. For. Meteorol. 2014, 189–190, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crout, N.; Kokkonen, T.; Jakeman, A.; Norton, J.; Newham, L.; Anderson, R.; Assaf, H.; Croke, B.; Gaber, N.; Gibbons, J.; et al. Chapter two good modelling practice. Dev. Integr. Environ. Assess. 2008, 3, 15–31. [Google Scholar]
- Camargo Rodriguez, A.V.; Ober, E.S. AquaCropR: Crop Growth Model for R. Agronomy 2019, 9, 378. [Google Scholar] [CrossRef] [Green Version]
- Agele, S.; Famuwagun, B.; Ogunleye, A. Effects of shade on microclimate, canopy characteristics and light integrals in dry season field-grown cocoa (Theobroma cacao L.) seedlings. J. Hortic. Sci. 2016, 11, 47–56. [Google Scholar]
- Soltani, A.; Sinclair, T.R. Modeling Physiology of Crop Development Growth and Yield; CABI: Wallingford, UK, 2012; pp. 1–8. [Google Scholar]
- Baracaldo, A.; Carvajal, R.; Romero, A.P.; Prieto, A.M.; Garcia, F.J.; Fischer, G.; Miranda, D. El anegamiento afecta el crecimiento y producción de biomasa en tomate chonto (Solanum lycopersicum L.), cultivado bajo sombrio. Rev. Colomb. Cienc. Hortic. 2014, 8, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, L.F.Q.; Castelblanco, S.G.; Jerez, A.G.; Guerrero, N.M. Caracterización de tres índices de cosecha de cacao de los clones CCN51, ICS60 e ICS 95, en la montaña santandereana, Colombia. Rev. Investig. Agrar. Ambient. 2015, 6, 252–265. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Johnstone, P.R.; Chakwizira, E.; Brown, H.E. Radiation capture and radiation use efficiency in response to N supply for crop species with contrasting canopies. Field Crops Res. 2013, 150, 126–134. [Google Scholar] [CrossRef]
- Bonhomme, R. Beware of comparing RUE values calculated from PAR vs solar radiation or absorbed vs intercepted radiation. Field Crops Res. 2000, 68, 247–252. [Google Scholar] [CrossRef]
- Danner, M.; Locherer, M.; Hank, T.; Richter, K. Measuring Leaf Area Index (LAI) with the LI-Cor LAI 2200C or LAI-2200 (+2200Clear Kit). Theory, measurement, problems, interpretation. In EnMAP Field Guide Technical Report; LI-Cor, Inc. Biosciences: Lincoln, NE, USA, 2015. [Google Scholar]
- Romero, A.P.; Alarcón, A.; Valbuena, R.I.; Galeano, C.H. Physiological assessment of water stress in potato using spectral information. Front. Plant Sci. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Viña, A.; Gitelson, A.A.; Nguy-Robertson, A.L.; Peng, Y. Comparison of different vegetation indices for the remote assessment of green leaf area index of crops. Remote Sens. Environ. 2011, 115, 3468–3478. [Google Scholar] [CrossRef]
- Amir, J.; Sinclair, T. A model of water limitation on spring wheat growth and yield. Field Crops Res. 1991, 28, 59–69. [Google Scholar] [CrossRef]
- Stöckle, C.O.; Donatelli, M.; Nelson, R. CropSyst, a cropping systems simulation model. Eur. J. Agron. 2003, 18, 289–307. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Raes, D.; Fereres, E. AquaCrop—The FAO Crop Model to Simulate Yield Response to Water: I. Concepts and Underlying Principles. Agron. J. 2009, 101, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Saunders, L.G. Methods for studying forcipomyia midges, with special reference to cacao-pollinating species (Diptera, ceratopogonidae). Can. J. Zool. 1959, 37, 33–51. [Google Scholar] [CrossRef]
- Kaufmann, T. Studies on the ecology and biology of a cocoa pollinator, Forcipomyia squamipennis L. (Diptera, Ceratopogonidae), in Ghana. Bull. Entomol. Res. 1975, 65, 263–268. [Google Scholar] [CrossRef]
- Sotomayor Chávez, A.C. Evaluación del riesgo ambiental de la aplicación de cuatro pesticidas sobre el polinizador Forcipomyia spp.(Díptera: Ceratopogonidae) del cultivo de cacao (Theobroma cacao L.), en San Martín, Perú. Ing. Ambient. 2020. Available online: https://hdl.handle.net/20.500.12805/1483 (accessed on 21 November 2021).
- Frimpong-Anin, K.; Adjaloo, M.K.; Kwapong, P.K.; Oduro, W. Structure and stability of cocoa flowers and their response to pollination. J. Bot. 2014. [Google Scholar] [CrossRef]
- Young, A.M. Seasonal differences in abundance and distribution of cocoa-pollinating midges in relation to flowering and fruit set between shaded and sunny habitats of the La Lola Cocoa Farm in Costa Rica. J. Appl. Ecol. 1983, 20, 801–831. [Google Scholar] [CrossRef]
- Young, A.M. Population Biology of Tropical Insects; Springer Science: Boston, MA, USA, 2012. [Google Scholar]
- Aneja, M.; Gianfagna, T.; Ng, E. The roles of abscisic acid and ethylene in the abscission and senescence of cocoa flowers. Plant Growth Regul. 1999, 27, 149–155. [Google Scholar] [CrossRef]
- Beyer Jr, E.M. Abscission: The initial effect of ethylene is in the leaf blade. Plant Physiol. 1975, 55, 322–327. [Google Scholar] [CrossRef]
- Frimpong, E.; Gordon, I.; Kwapong, P.; Gemmill-Herren, B. Dynamics of cocoa pollination: Tools and applications for surveying and monitoring cocoa pollinators. Int. J. Trop. Insect Sci. 2009, 29, 62–69. [Google Scholar] [CrossRef]
- Frimpong, E.A.; Gemmill-Herren, B.; Gordon, I.; Kwapong, P.K. Dynamics of insect pollinators as influenced by cocoa production systems in Ghana. J. Pollinat. Ecol. 2011, 5, 74–80. [Google Scholar] [CrossRef]
- Vaughton, G.; Ramsey, M. Pollinators and seed production. In Seed development and germination; Routledge: London, UK, 2017; pp. 475–490. [Google Scholar]
- Alvim, P.d.T.; Machado, A.; Vello, F. Physiological responses of cacao to environmental factors. Rev. Theobroma 1974, 4, 3–25. [Google Scholar]
- End, M.J. A Study of the Effects of the Photo-Thermal Environment on Fruit and Seed Growth and Development in Theobroma cacao L.; The University of Reading: Reading, UK, 1991. [Google Scholar]
- Osorio, S.; Scossa, F.; Fernie, A. Molecular regulation of fruit ripening. Front. Plant Sci. 2013, 4, 198. [Google Scholar] [CrossRef] [Green Version]
- Raymundo, R.; Asseng, S.; Prassad, R.; Kleinwechter, U.; Concha, J.; Condori, B.; Bowen, W.; Wolf, J.; Olesen, J.E.; Dong, Q.; et al. Performance of the SUBSTOR-potato model across contrasting growing conditions. Field Crops Res. 2017, 202, 57–76. [Google Scholar] [CrossRef] [Green Version]
- Confalonieri, R.; Orlando, F.; Paleari, L.; Stella, T.; Gilardelli, C.; Movedi, E.; Pagani, V.; Cappelli, G.; Vertemara, A.; Alberti, L.; et al. Uncertainty in crop model predictions: What is the role of users? Environ. Model. Softw. 2016, 81, 165–173. [Google Scholar] [CrossRef]
- Deblonde, P.; Ledent, J. Effects of moderate drought conditions on green leaf number, stem height, leaf length and tuber yield of potato cultivars. Eur. J. Agron. 2001, 14, 31–41. [Google Scholar] [CrossRef]
- Karunaratne, A.; Azam-Ali, S.; Al-Shareef, I.; Sesay, A.; Jørgensen, S.; Crout, N. Modelling the canopy development of bambara groundnut. Agric. For. Meteorol. 2010, 150, 1007–1015. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Tsum | RUE | Yield * |
|---|---|---|---|
| Apartado | 2906 | 0.6 | 3378 |
| Arauca | 2764 | 0.7 | 3981 |
| Santander | 2016 | 0.6 | 2687 |
| Cali | 1912 | 0.5 | 1900 |
| Caldas | 1192 | 0.6 | 740 |
| File | Variable Name | Value |
|---|---|---|
| SoilName | Loamy sand4 | |
| InitialFsolar | 0.01 | |
| Treatment | Weather | KOKO (.WTH file name) |
| CO | 400 ppm | |
| SowingDate | Flowering Date (FD) | |
| Crop cycle DAP | 200 days | |
| LAI | 1.8 | |
| Observation | FSolar | 0.70 |
| Biomass | 40 kg dry mass per plant | |
| Harvest index | 0.3 | |
| Cultivar | 150A | 680 C day |
| 150B | 680 C day | |
| Tbase | 10 C | |
| Topti | 26 C | |
| Species | MaxT | 35 C |
| ExtremeT | 40 C | |
| CORUE | 0.09 C | |
| S-water | 0 ARID index |
| Region | Apartado | Arauca | Santander | Cali | Caldas | Overal |
|---|---|---|---|---|---|---|
| RMMSE% | 3 | 6.05 | 10.06 | 8.5 | 14.90 | 7.2 |
| Month | Santander | Arauca | Cali | Apartado | Caldas |
|---|---|---|---|---|---|
| January | 166.5 | 166.5 | 169.5 | 171.5 | 158.5 |
| February | 166 | 171 | 171 | 173 | 163 |
| March | 165 | 171 | 172 | 176 | 167 |
| April | 165.5 | 172.5 | 173 | 178.5 | 170 |
| May | 166.5 | 174.5 | 174.5 | 181 | 175 |
| June | 169 | 173.5 | 177 | 182 | 175.5 |
| July | 176 | 191 | 175 | 179 | 173.5 |
| August | 181 | 186 | 175 | 178.5 | 171 |
| September | 176 | 182.5 | 175 | 176.5 | 168 |
| October | 179 | 176 | 172.5 | 173 | 163.5 |
| November | 173.5 | 170 | 170.5 | 171.5 | 159.5 |
| December | 169 | 166 | 168.5 | 171 | 157 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero Vergel, A.P.; Camargo Rodriguez, A.V.; Ramirez, O.D.; Arenas Velilla, P.A.; Gallego, A.M. A Crop Modelling Strategy to Improve Cacao Quality and Productivity. Plants 2022, 11, 157. https://doi.org/10.3390/plants11020157
Romero Vergel AP, Camargo Rodriguez AV, Ramirez OD, Arenas Velilla PA, Gallego AM. A Crop Modelling Strategy to Improve Cacao Quality and Productivity. Plants. 2022; 11(2):157. https://doi.org/10.3390/plants11020157
Chicago/Turabian StyleRomero Vergel, Angela Patricia, Anyela Valentina Camargo Rodriguez, Oscar Dario Ramirez, Paula Andrea Arenas Velilla, and Adriana Maria Gallego. 2022. "A Crop Modelling Strategy to Improve Cacao Quality and Productivity" Plants 11, no. 2: 157. https://doi.org/10.3390/plants11020157
APA StyleRomero Vergel, A. P., Camargo Rodriguez, A. V., Ramirez, O. D., Arenas Velilla, P. A., & Gallego, A. M. (2022). A Crop Modelling Strategy to Improve Cacao Quality and Productivity. Plants, 11(2), 157. https://doi.org/10.3390/plants11020157