
Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Material
2.1.1. Field Grown Maize for Metabolite Quantification
2.1.2. Greenhouse Grown Maize for Carbohydrate Quantification and RNA Sequencing
2.2. Iodine Staining
2.3. Non-Structural Carbohydrate Quantification
2.4. RNA Extraction
2.5. RNAseq Analysis
2.6. Differential Gene Expression Analysis
2.7. GO Enrichment Analysis
2.8. Quantitative Real-Time PCR (qRT-PCR)
2.9. Gene Expression of Sucrose-Starch Genes in B73 Genome Atlas
2.10. Statistical Analysis
3. Results
3.1. Hexose to Sucrose Ratio Increases at Early Reproductive Stages
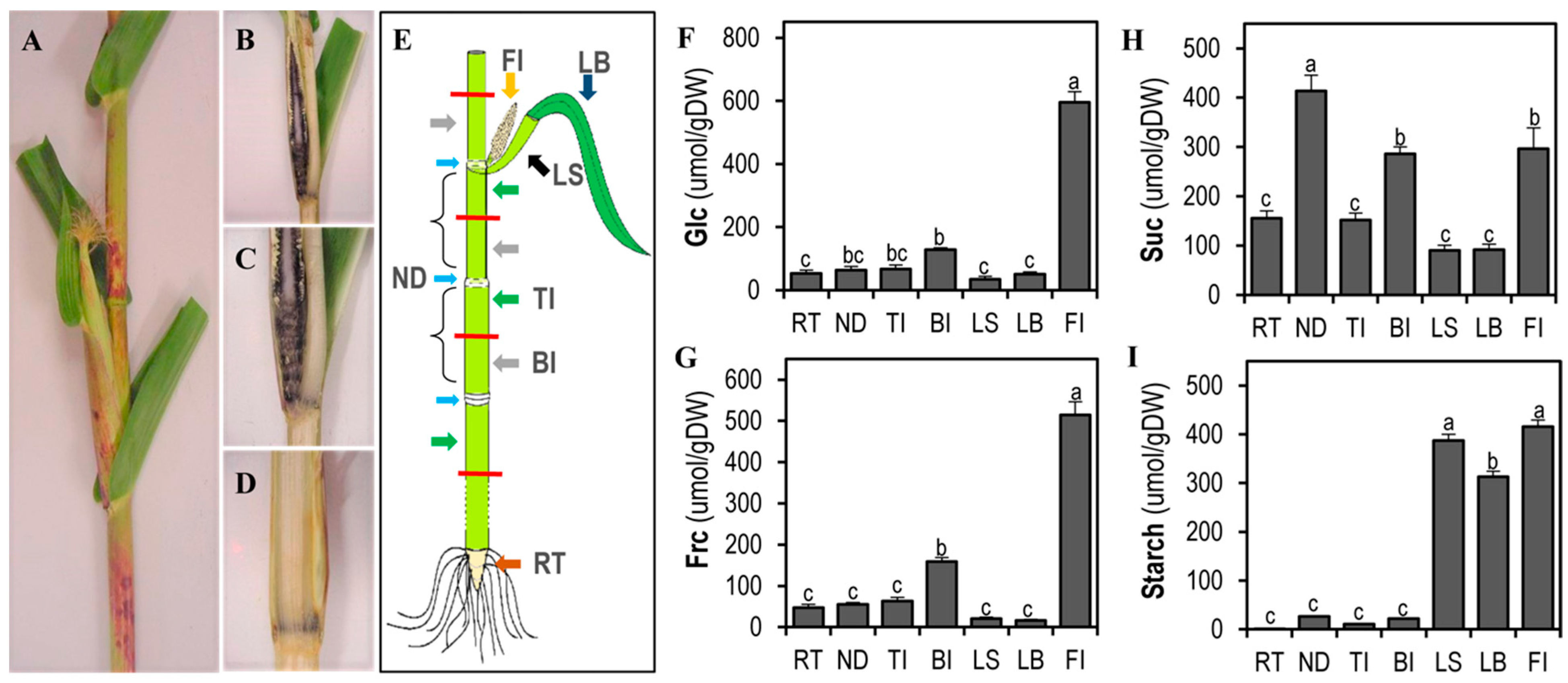
3.2. The Female Inflorescence before Pollination Has High Sink Strength
3.3. Differential Gene Expression between the Female Inflorescence and the Top Internode of the Stem
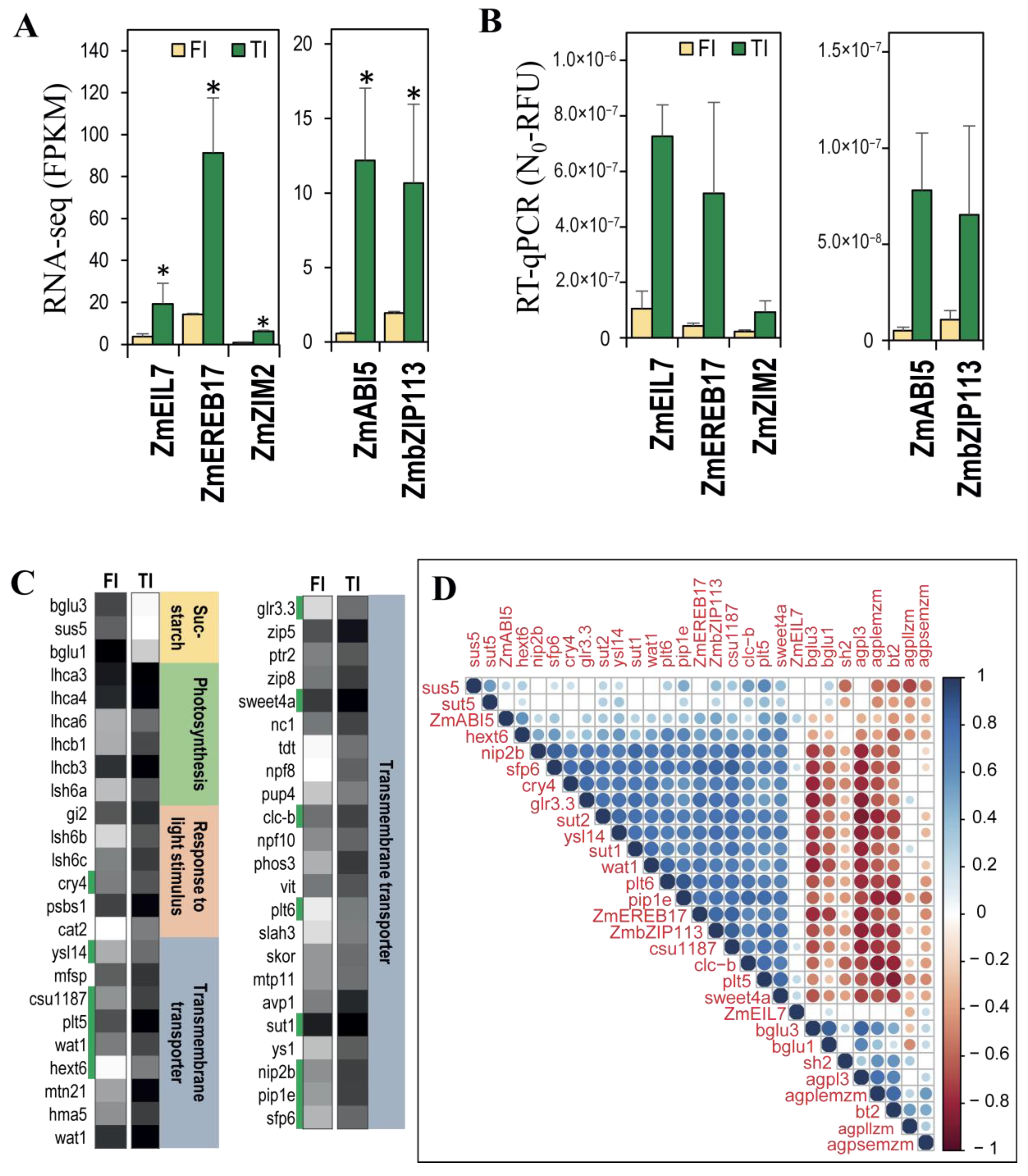
3.4. qRT-PCR Validation of Gene Expression for Sucrose-Starch Metabolism Enzymes
3.5. The Expression of the bZip113 and Ereb17 Transcription Factor Genes Was Highly Correlated with the Expression of Sucrose Transporters
4. Discussion
4.1. Starch Accumulation in Maize Physiology
4.2. The Function of The Maize Stem in Carbon Partitioning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slewinski, T.L.; Braun, D.M. Current Perspectives on the Regulation of Whole-Plant Carbohydrate Partitioning. Plant Sci. 2010, 178, 341–349. [Google Scholar] [CrossRef]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How Can We Make Plants Grow Faster? A Source-Sink Perspective on Growth Rate. J. Exp. Bot. 2016, 67, 31–45. [Google Scholar] [CrossRef]
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-Sink Relationships in Crop Plants and Their Influence on Yield Development and Nutritional Quality. Front. Plant Sci. 2018, 871, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gambín, B.L.; Borrás, L.; Otegui, M.E. Source-Sink Relations and Kernel Weight Differences in Maize Temperate Hybrids. Field Crop. Res. 2006, 95, 316–326. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Bush, D.R. Carbohydrate Export from the Leaf: A Highly Regulated Process and Target to Enhance Photosynthesis and Productivity. Plant Physiol. 2011, 155, 64–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lu, Q.; Wen, X.; Lu, C. Enhanced Sucrose Loading Improves Rice Yield by Increasing Grain Size. Plant Physiol. 2015, 169, 2848–2862. [Google Scholar] [CrossRef] [Green Version]
- Bancal, P. Source-Sink Partitioning. Do We Need Munch? J. Exp. Bot. 2002, 53, 1919–1928. [Google Scholar] [CrossRef]
- Streb, S.; Zeeman, S.C. Starch Metabolism in Arabidopsis. Arab. Book 2012, 10, e0160. [Google Scholar] [CrossRef] [Green Version]
- Van Harsselaar, J.K.; Lorenz, J.; Senning, M.; Sonnewald, U.; Sonnewald, S. Genome-Wide Analysis of Starch Metabolism Genes in Potato (Solanum tuberosum L.). BMC 2017, 18, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stitt, M.; Zeeman, S.C. Starch Turnover: Pathways, Regulation and Role in Growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Baroja-Fernández, E.; Bahaji, A.; Muñoz, F.J.; Ovecka, M.; Montero, M.; Sesma, M.T.; Alonso-Casajús, N.; Almagro, G.; Sánchez-López, A.M.; et al. Enhancing Sucrose Synthase Activity Results in Increased Levels of Starch and Adp-Glucose in Maize (Zea mays L.) Seed Endosperms. Plant Cell Physiol. 2013, 54, 282–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennen-Bierwagen, T.A.; Lin, Q.; Grimaud, F.; Planchot, V.; Keeling, P.L.; James, M.G.; Myers, A.M. Proteins from Multiple Metabolic Pathways Associate with Starch Biosynthetic Enzymes in High Molecular Weight Complexes: A Model for Regulation of Carbon Allocation in Maize Amyloplasts. Plant Physiol. 2009, 149, 1541–1559. [Google Scholar] [CrossRef] [Green Version]
- Tomasik, P.; Horton, D. Enzymatic Conversions of Starch, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 68, ISBN 9780123965233. [Google Scholar]
- Pfister, B.; Zeeman, S.C. Formation of Starch in Plant Cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smidansky, E.D.; Clancy, M.; Meyer, F.D.; Lanning, S.P.; Blake, N.K.; Talbert, L.E.; Giroux, M.J. Enhanced ADP-Glucose Pyrophosphorylase Activity in Wheat Endosperm Increases Seed Yield. Proc. Natl. Acad. Sci. USA 2002, 99, 1724–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smidansky, E.D.; Martin, J.M.; Hannah, L.C.; Fischer, A.M.; Giroux, M.J. Seed Yield and Plant Biomass Increases in Rice Are Conferred by Deregulation of Endosperm ADP-Glucose Pyrophosphorylase. Planta 2003, 216, 656–664. [Google Scholar] [CrossRef]
- Kang, G.; Liu, G.; Peng, X.; Wei, L.; Wang, C.; Zhu, Y.; Ma, Y.; Jiang, Y.; Guo, T. Increasing the Starch Content and Grain Weight of Common Wheat by Overexpression of the Cytosolic AGPase Large Subunit Gene. Plant Physiol. Biochem. 2013, 73, 93–98. [Google Scholar] [CrossRef]
- Mugford, S.T.; Fernandez, O.; Brinton, J.; Flis, A.; Krohn, N.; Encke, B.; Feil, R.; Sulpice, R.; Lunn, J.E.; Stitt, M.; et al. Regulatory Properties of ADP Glucose Pyrophosphorylase Are Required for Adjustment of Leaf Starch Synthesis in Different Photoperiods. Plant Physiol. 2014, 166, 1733–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-González, C.; Juárez-Colunga, S.; Morales-Elías, N.C.; Tiessen, A. Exploring Regulatory Networks in Plants: Transcription Factors of Starch Metabolism. PeerJ 2019, 2019, 1–27. [Google Scholar] [CrossRef]
- Chen, J.; Yi, Q.; Cao, Y.; Wei, B.; Zheng, L.; Xiao, Q.; Xie, Y.; Gu, Y.; Li, Y.; Huang, H.; et al. ZmbZIP91 Regulates Expression of Starch Synthesis-Related Genes by Binding to ACTCAT Elements in Their Promoters. J. Exp. Bot. 2016, 67, 1327–1338. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Xie, S.; Xiao, Q.; Wei, B.; Zheng, L.; Wang, Y.; Cao, Y.; Zhang, X.; Long, T.; Li, Y.; et al. Sucrose and ABA Regulate Starch Biosynthesis in Maize through a Novel Transcription Factor, ZmEREB156. Sci. Rep. 2016, 6, 27590. [Google Scholar] [CrossRef]
- Wang, J.-C.; Xu, H.; Zhu, Y.; Liu, Q.-Q.; Cai, X.-L. OsbZIP58, a Basic Leucine Zipper Transcription Factor, Regulates Starch Biosynthesis in Rice Endosperm. J. Exp. Bot. 2013, 64, 3453–3466. [Google Scholar] [CrossRef] [PubMed]
- Fukayama, H.; Morita, R.; Sugino, M.; Hatanaka, T.; Misoo, S. CO2 Responsive CCT Protein, CRCT Is a Positive Regulator of Starch Synthesis in Vegetative Organs of Rice. Plant Physiol. 2015, 167, 1321–1331. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.M.; Lo, S.F.; Ho, T.H.D. Source-Sink Communication: Regulated by Hormone, Nutrient, and Stress Cross-Signaling. Trends Plant Sci. 2015, 20, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Angeles-Núñez, J.G.; Tiessen, A. Arabidopsis Sucrose Synthase 2 and 3 Modulate Metabolic Homeostasis and Direct Carbon towards Starch Synthesis in Developing Seeds. Planta 2010, 232, 701–718. [Google Scholar] [CrossRef]
- Vargas-Ortiz, E.; Espitia-Rangel, E.; Tiessen, A.; Délano-Frier, J.P. Grain Amaranths Are Defoliation Tolerant Crop Species Capable of Utilizing Stem and Root Carbohydrate Reserves to Sustain Vegetative and Reproductive Growth after Leaf Loss. PLoS ONE 2013, 8, e67879. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering Splice Junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential Gene and Transcript Expression Analysis of RNA-Seq Experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.G.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An Empirical Bayes Hierarchical Model for Inference in RNA-Seq Experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape Plug-in to Decipher Functionally Grouped Gene Ontology and Pathway Annotation Networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat Genet 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER Version 14: More Genomes, a New PANTHER GO-Slim and Improvements in Enrichment Analysis Tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Colunga, S.; López-González, C.; Morales-Elías, N.C.; Massange-Sánchez, J.A.; Trachsel, S.; Tiessen, A. Genome-Wide Analysis of the Invertase Gene Family from Maize. Plant Mol. Biol. 2018, 97, 385–406. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F. Assumption-Free Analysis of Quantitative Real-Time Polymerase Chain Reaction (PCR) Data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; Van Den Hoff, M.J.B.; Moorman, A.F.M. Amplification Efficiency: Linking Baseline and Bias in the Analysis of Quantitative PCR Data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]
- Massange-Sánchez, J.A.; Casados-Vázquez, L.E.; Juarez-Colunga, S.; Sawers, R.J.H.; Tiessen, A. The Phosphoglycerate Kinase (Pgk) Gene Family of Maize (Zea mays Var. B73). Plants 2020, 9, 1639. [Google Scholar] [CrossRef]
- Yilmaz, A.; Nishiyama, M.Y.; Fuentes, B.G.; Souza, G.M.; Janies, D.; Gray, J.; Grotewold, E. GRASSIUS: A Platform for Comparative Regulatory Genomics across the Grasses. Plant Physiol. 2009, 149, 171–180. [Google Scholar] [CrossRef] [Green Version]
- MacNeill, G.J.; Mehrpouyan, S.; Minow, M.A.A.; Patterson, J.A.; Tetlow, I.J.; Emes, M.J. Starch as a Source, Starch as a Sink: The Bifunctional Role of Starch in Carbon Allocation. J. Exp. Bot. 2017, 68, 4433–4453. [Google Scholar] [CrossRef]
- Shane, M.W.; McCully, M.E.; Canny, M.J. The Vascular System of Maize Stems Revisited: Implications for Water Transport and Xylem Safety. Ann. Bot. 2000, 86, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Scofield, G.N.; Ruuska, S.A.; Aoki, N.; Lewis, D.C.; Tabe, L.M.; Jenkins, C.L.D. Starch Storage in the Stems of Wheat Plants: Localization and Temporal Changes. Ann. Bot. 2009, 103, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salleo, S.; Trifilò, P.; Esposito, S.; Nardini, A.; Lo Gullo, M.A. Starch-to-Sugar Conversion in Wood Parenchyma of Field-Growing Laurus Nobilis Plants: A Component of the Signal Pathway for Embolism Repair? Funct. Plant Biol. 2009, 36, 815. [Google Scholar] [CrossRef] [PubMed]
- Bennetzen, J.; Hake, S. Handbook of Maize, Its Biology; Springer: New York, NY, USA, 2009; ISBN 9780387794174. [Google Scholar]
- Reeves, W.M.; Lynch, T.J.; Mobin, R.; Finkelstein, R.R. Direct Targets of the Transcription Factors ABA-Insensitive(ABI)4 and ABI5 Reveal Synergistic Action by ABI4 and Several BZIP ABA Response Factors. Plant Mol. Biol. 2011, 75, 347–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rook, F.; Hadingham, S.A.; Li, Y.; Bevan, M.W. Sugar and ABA Response Pathways and the Control of Gene Expression. Plant, Cell Environ. 2006, 29, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Kerchev, P.I.; Hancock, R.D. The ABA-INSENSITIVE-4 (ABI4) Transcription Factor Links Redox, Hormone and Sugar Signaling Pathways. Plant Signal. Behav. 2012, 7, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Weise, S.E.; van Wijk, K.J.; Sharkey, T.D. The Role of Transitory Starch in C3, CAM, and C4 Metabolism and Opportunities for Engineering Leaf Starch Accumulation. J. Exp. Bot. 2011, 62, 3109–3118. [Google Scholar] [CrossRef]
- Lloyd, J.R.; Kossmann, J. Transitory and Storage Starch Metabolism: Two Sides of the Same Coin? Curr. Opin. Biotechnol. 2015, 32, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Nagler, M.; Nukarinen, E.; Weckwerth, W.; Nägele, T. Integrative Molecular Profiling Indicates a Central Role of Transitory Starch Breakdown in Establishing a Stable C/N Homeostasis during Cold Acclimation in Two Natural Accessions of Arabidopsis Thaliana. BMC Plant Biol. 2015, 15, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Scialdone, A.; Howard, M. How Plants Manage Food Reserves at Night: Quantitative Models and Open Questions. Front. Plant Sci. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, C.A.; Paul, M.J.; Foyer, C.H. Metabolite Transport and Associated Sugar Signalling Systems Underpinning Source/Sink Interactions. Biochim. Biophys. Acta-Bioenerg. 2016, 1857, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Bel, A.J.E.; Hess, P.H. Hexoses as Phloem Transport Sugars: The End of a Dogma? J. Exp. Bot. 2008, 59, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.P.M.; Bräutigam, A. The Role of Membrane Transport in Metabolic Engineering of Plant Primary Metabolism. Curr. Opin. Biotechnol. 2013, 24, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.M.; Wang, L.; Ruan, Y.-L. Understanding and Manipulating Sucrose Phloem Loading, Unloading, Metabolism, and Signalling to Enhance Crop Yield and Food Security. J. Exp. Bot. 2014, 65, 1713–1735. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-González, C.; Juárez-Colunga, S.; Trachsel, S.; Marsch-Martínez, N.; Gillmor, C.S.; Tiessen, A. Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose. Plants 2022, 11, 238. https://doi.org/10.3390/plants11030238
López-González C, Juárez-Colunga S, Trachsel S, Marsch-Martínez N, Gillmor CS, Tiessen A. Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose. Plants. 2022; 11(3):238. https://doi.org/10.3390/plants11030238
Chicago/Turabian StyleLópez-González, Cristal, Sheila Juárez-Colunga, Samuel Trachsel, Nayelli Marsch-Martínez, C. Stewart Gillmor, and Axel Tiessen. 2022. "Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose" Plants 11, no. 3: 238. https://doi.org/10.3390/plants11030238
APA StyleLópez-González, C., Juárez-Colunga, S., Trachsel, S., Marsch-Martínez, N., Gillmor, C. S., & Tiessen, A. (2022). Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose. Plants, 11(3), 238. https://doi.org/10.3390/plants11030238