1. Introduction
In recent decades, there has been a strong demand to gradually replace intensive agriculture with sustainable cultivation practices, which are based on the use of organic amendments that are able to preserve the integrity of the soil and the healthiness of crops [
1,
2,
3]. Plants need not only light and water for their metabolism, but also a complete mineral nutrition, whose availability is governed by soil properties and the efficiency of water uptake by the roots. Currently, the functionality and productivity of the soil are seriously compromised by the intensive use for crops and by the degradation of fertile soils [
4]; the use of biofertilizers in agriculture help plants to enhance their absorption of nutrient elements from the soil and improve their growth [
5]. The production of organic fertilizers from the urban organic waste is expected to increase, and the challenge at the moment is to tackle these problems together by transforming food waste into a sustainable resource to enrich the soils and enhance their nutritional quality and regeneration. Among the waste sources, those of the food residues and the municipal biosolids seem to be the most promising in the urban sustainable recycling process [
6]. Unfortunately, current recycling processes take months to degrade organic wastes, thus representing the main limiting factor for a large-scale application. Alternative processes, based on waste drying or high-temperature aerobic fermentation were advanced to overcome this limitation [
7,
8]. Nowadays, the classical Fenton reaction [
9] is considered as one of a set of advanced oxidation processes (AOPs) and is widely used for the chemical treatment of wastewater, industrial sludge, landfill leachate, soils, and sediments, which are contaminated with biorefractory organic compounds including phenols, dyes, pesticides, organic solvents, pharmaceuticals, and domestic chemicals [
10]. Specifically, the fast degradation of urban solid waste using the Fenton reaction has already been achieved [
11]; the authors reported that after a few hours of incubation, the Fe (II) ion catalytic oxidation pathway was able to degrade dry organic matrices with a C/N ratio lower than 12, with a high degree of oxidative decomposition converted into low-molecular-weight compounds at a high-oxidation state.
The aim of this research was to assess the effects of the soil amendment formulation reported as the fast-composted soil amendment (FCA), obtained by urban, organic solid waste fractions following the Fenton composting reaction on the growth of Lactuca sativa L. seedlings. Lettuce is one of the most intensively cultivated vegetables in Mediterranean countries. Millions of tons of lettuce for fresh consumption are produced annually in the EU, mainly in Mediterranean countries (FAO, 2022). Intensive lettuce cultivation practices are large consumers of fertilizers, thus entailing significant costs in terms of energy and the environment. Therefore, providing alternatives that improve the sustainability of these agricultural ecosystems without reducing productivity is of great interest. Under our experimental conditions, L. sativa was grown in a short-day period to prevent flowering and thus evaluates plant growth in its vegetative phase. Morphological, biochemical, and molecular analyzes have been performed to elucidate the metabolic behavior of the leaves and roots of plants grown on soil enriched with the FCA.
3. Discussion
Application of organic fertilizers can significantly improve the growth, yield, and quality of the lettuce without negatively impacting the environment and human health [
12]. Authors reported that, when composted urban waste was used, there were inhibitory or growth-inducing effects of the lettuce seedlings, possibly due to a combination of the high electrical conductivity, ammonia toxicity, and degree of stabilization of this compost [
13,
14]. Further investigations on the effects of the types of organic amendments on lettuce seedlings reported an enhanced plant biomass, and a higher content of the Rubisco large subunit and soluble proteins; on the contrary, an antagonistic effect was observed on the chlorophyll content [
12].
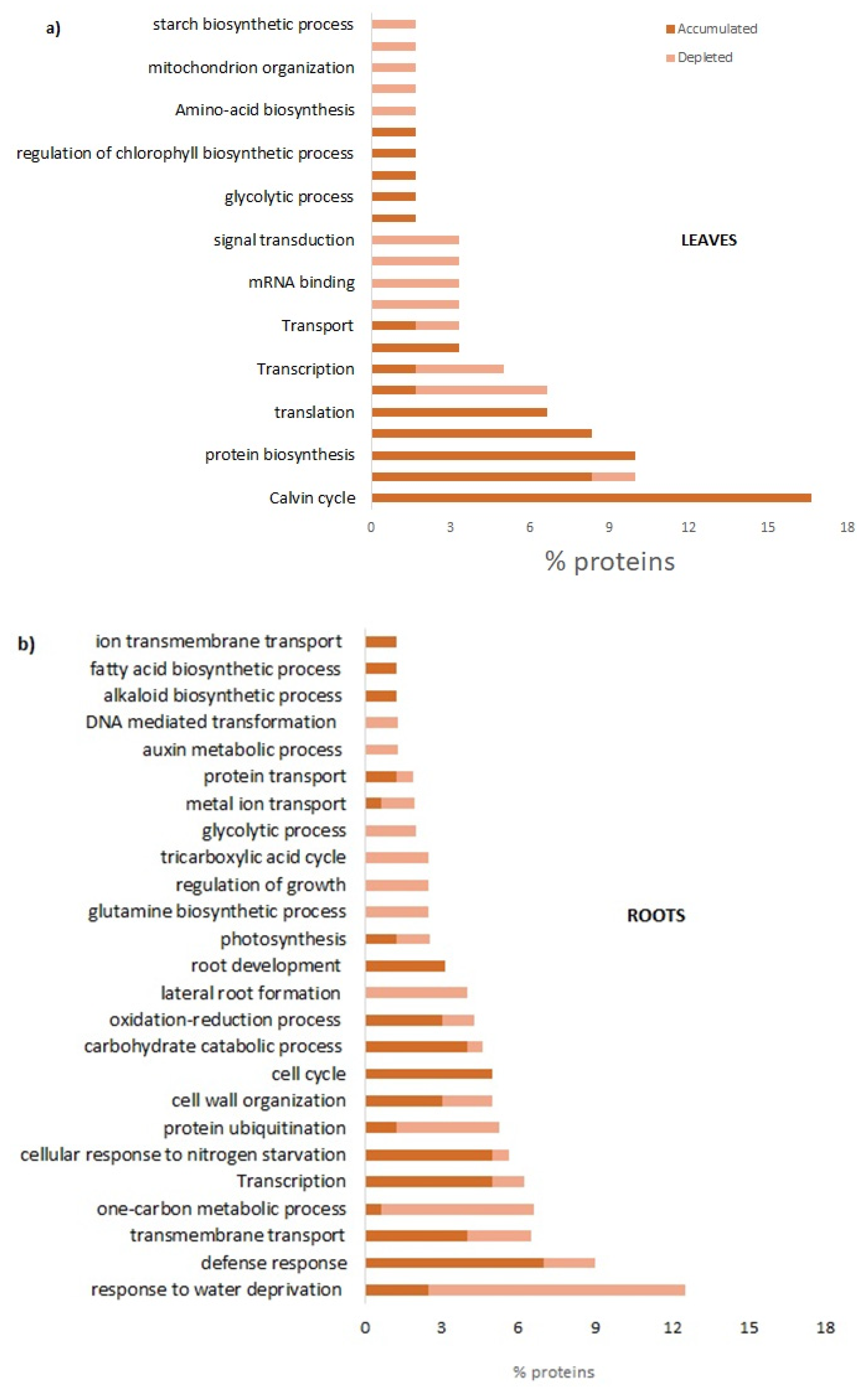
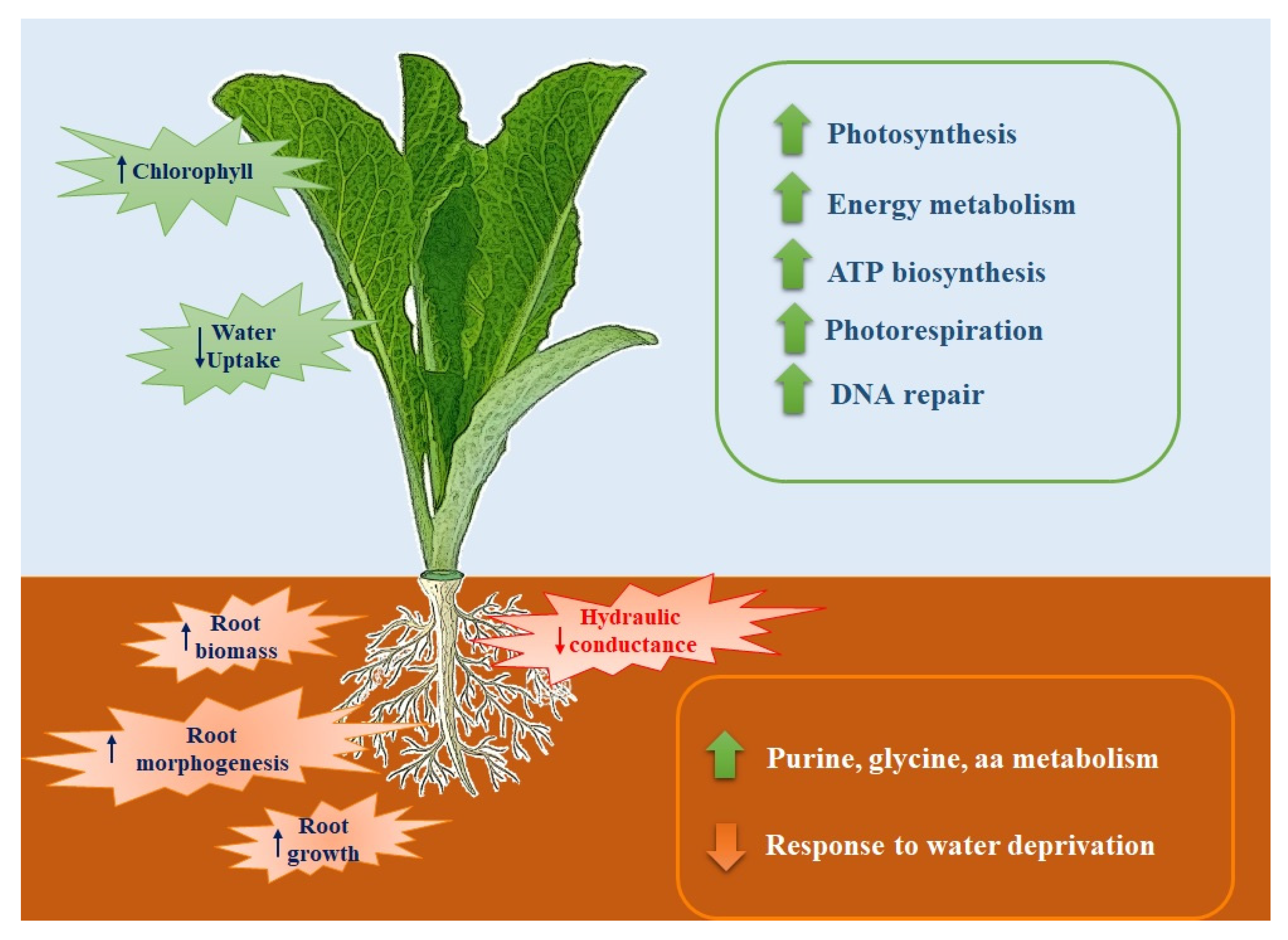
Fast-composted soil amendment (FCA) produced by the urban, organic solid waste fractions and stabilized by the Fenton’s reaction procedure, revealed after 8 h of treatment, a good degree of oxidative decomposition with C/N ratios of 22, compared to an initial value of 26. In addition, the FCA exhibited a dark-brown color, was odorless, and had a humidity of 15%. The product cannot be considered as a conventional compost, but claimed to be a stabilized organic matrix. In this case, the material does not go, at least within 6 months, in further biochemical transformation when stored under dry conditions. Property of the material as a soil fertilizer was assessed on Lactuca sativa seedlings that underwent modification of the biochemical and molecular behavior; the FCA treatment, In fact, positively affected both root growth and photosynthesis. After 28 days treatment of L. sativa seedlings, the root biomass and chlorophyll content were found to have significantly increased with respect to the control seedlings. The chlorophyll physiological adjustments induced by the FCA treatment appeared to be linked to an increase in photosynthesis, as the proteins related to this metabolism accumulated in the shoots. Under our conditions, however, the shoot biomass was not affected by the treatment, indicating that the amendment exerted contrasting effects on shoot metabolism.
The FCA treatment also promoted primary and secondary metabolism in the leaves; the mitotic intra-S DNA damage checkpoint signaling process was enriched seventy times with respect to the untreated leaves; during the S-phase of the cell cycle, and in the event of problems during the replication process, DNA integrity checkpoints are activated slowing down the cell cycle to grant the cell time to repair the damage [
15]; evidence has also suggested that this well-known mechanism has a role as a growth regulator processer in plants [
16]. On this view, the FCA imposed a stress condition that, however, the leaf cells coped with by activating the mechanisms that slow down the mitosis to repair the DNA, this might lead to a slowdown in tissue growth.
Amendment also induced the enrichment of the S-adenosylmethionine (SAM) cycle; SAM is a key enzyme involved in many important biological processes, such as ethylene and polyamine biosynthesis, transmethylation, and transulfuration; SAM genes showed differential expression in response to abiotic stresses and exogenous hormone treatments [
17]. Polyamines, for example, are important hormones that regulate cell growth during stress responses, pollen and flower development, and the protection of photosystem II [
18]. The cell demand for SAM compounds, of course, may change markedly under different growth conditions with a metabolic cost of ATP consumption. In this regard, the metabolic processes of ATP production, mainly through photophosphorylation, have been enriched to cope with the high energy demand under treatment with FCA; the energy metabolism that uses carbohydrates was induced through the processes of photosynthetic gluconeogenesis, the biosynthesis of sucrose and glycolysis. To complete the effects on the metabolic patterns induced by the FCA treatment, the upregulation of processes involving the biosynthesis and metabolisms of glycine have been observed. It is well known that endogenous glycine accumulation mediates abiotic stress tolerance in plants involving the osmotic regulation [
19]. In response to the FCA treatment, glycine should be synthesized in excess to adjust the osmotic stress induced by stress to maintain the sub-cellular structures and reduce the oxidative damage. In addition, the catabolic process of amino acids and dicarboxylic acids could be related to the high energy demand of treated leaves; catabolic pathways for several amino acids including alanine or glutamine are very short, and they can be directly converted to pyruvate by alanine aminotransferases and to glutamate, respectively, contributing substantially to the energy state of plant cells under certain physiological conditions [
20].
In roots, the FCA treatment strongly enriched the organs morphogenesis and development; root cap development, lateral root formation, and post-embryonic root morphogenesis were the main biological processes enriched by the treatment. It is well known that morphogenetic processes are the basis of new organ formation; lateral roots morphogenesis is a decisive process during root system formation [
21]. Regarding the root cap, it is the terminal tissue of the root of most plants. Historical evidence has shown that the root cap has not only the role of protecting proximal root meristem, but also to direct root growth in response to stimuli such as gravity, light, gradients in temperature, humidity, ions, and other chemicals [
22]. Responses to water deprivation were strongly inhibited by the FCA treatment. The highly conserved plant aquaporins, known as plasma membrane intrinsic Proteins (PIPs), are the main gateways for cell membrane water exchange; in Arabidopsis, the inhibition of aquaporin expression in roots induced an increase in root growth [
23]. Researchers have found that if plants lack aquaporins, thus having an increased resistance for the water movement in leaf and root cells, they compensate for this effect by increasing their root surface; this finding completes the evidence that FCA strongly alters the plant water transport and root growth dynamic. Additionally, the biosynthetic processes of amino acids, the biosynthesis of ribonucleotides, and the biosynthetic process of carboxylic acids have also been enriched. Morphological, biochemical, and molecular findings were, then, found to be completely consistent with the significant root growth promotion of lettuce seedlings cultivated on FCA-amended soil. FCA induced, in fact, the enrichment of the primary metabolism of purine, amino acids, and carbohydrates. Biological processes of the S-adenosylmethionine (SAM) cycle were strongly enriched. SAM, as reported above, is the main methyl group donor useful for the methylation of DNA, RNA, protein, lignin, and flavonoids, and it also plays important roles in regulating plant development under both abiotic or biotic stress [
17], and heavy metal tolerance [
24].
4. Materials and Methods
4.1. Fast-Composted Soil Amendment (FCA) Production
The fast-composted soil amendment (FCA) was produced starting from samples of municipal solid waste. Waste was dried under vacuum and finely ground; 200 gr of dried powder were placed in the glass reactor, Fe
2+ (as the catalyst) at concentrations of 0.01% FeSO
4 (VEBI Istituto Biochimico s.r.l., Borgoricco (PD), Italy) and 0.6 × 10
−3 H
2O
2 (as the oxidant) (Panreac Applichem, Barcelona, Spain) were then added. The Fenton’s reaction was performed at pH 3.0, 60 °C, under a pressure of 0.96 bar for 8 h, during which the processes of chemical stabilization of the biomass took place. Further details of the reaction are reported in Roccotelli [
25]. For the elemental analysis of FCA, the stabilized biomasses were subjected to high-temperature combustion (T ≥ 900 °C) in a bomb containing oxygen under pressure and following the method described in Nelms [
26]; residues were analyzed by ICP/MS iCapQ (Thermofischer, Waltham, MA, USA) to evaluate the concentrations of C, H, and N in terms of their relative percentages. Results of the elemental analysis are reported in
Supplementary Table S3.
4.2. Plant Acclimation, Treatments, and Growth
Seedlings of
Latuca sativa L. var. longifolia (
n = 70), with four leaves each, were transplanted in a soft and porous commercial soil, (Humifly, Humiflora, Italy) and mixed with pine bark, light peat, and volcanic lapilli in a proportion of 2:1 (
v/
v). Seedlings of
L. sativa were acclimated under controlled temperatures and a short-day 10/14 h light/dark cycle. After seven days, each seedling was weighed and measured in length, and were then divided into three sub-cultivations of 35 seedlings each on soils whose compositions were as follows: (i) soil without amendment (control), and (ii) soil with 2% FCA [
27]. After 30 days cultivation, all seedlings were collected; for each sub-cultivation, whole plant weight, weight of the roots and leaves, number of total leaves for each seedling, and the number of healthy leaves were measured. The leaves and roots were, then frozen in liquid nitrogen and stored at −80 °C to be used for the physiological and molecular analyzes.
4.3. Chlorophyll Extraction and Measurement
A total of 1.0 g of frozen leaf tissue was ground in liquid nitrogen in a mortar to obtain a fine powder; 5 mL of 80% cold acetone was added to the tissue powder and incubated at 4 °C for 3 h under weak shaking; then the samples were centrifuged at 800×
g for 15 min. Then, 1 mL of crude supernatant was transferred in a cuvette and the absorbance was measured at 663 nm and 645 nm with the 7310 Jenway spectrophotometer. The concentrations of chlorophylls a and b, and of the total chlorophyll were determined by the following equations [
28]:
where chlorophylls content as mg/g fresh tissue were measured in six biological replicates.
4.4. Protein Extraction and Purification
4.4.1. Protein Extraction and Purification from Leaves
Proteins from the leaves of three biological replicates of the control and FCA treatments were extracted by the multistep procedures [
29]. For each extraction, 1.4 g of tissues were crushed in a mortar in liquid nitrogen until a fine powder was obtained. Ground plant tissue was homogenized with a volume of 10% TCA in acetone and centrifuged at 14,000×
g for 5 min; a volume of 10% TCA in water was added and centrifuged at 14,000×
g for 5 min. Subsequently, four washes were performed using 80% acetone in water. After centrifugation, the pellet containing the precipitated proteins was dried at room temperature. Approximately 100 mg of powdered tissue was dissolved in 0.8 mL of phenol (buffered with Tris HCL, pH8.0, Sigma, St. Louis, MO, USA) and 0.8 mL of SDS buffer (30% sucrose, 2% SDS, 0.1M Tris-HCl, pH8.0, 5% 2-mercaptoethanol) in a 2 mL microfuge tube. The samples were vortexed for 30 s and centrifuged at 14,000×
g for 5 min to allow proteins to solubilize in the phenol phase. The phenol phase was mixed with five volumes of 0.1 M ammonium acetate in cold methanol, and then the mixture was stored at −20 °C for 30 min to precipitate the proteins. Proteins were collected by centrifugation at 14,000×
g for 5 min. Two washes were then performed with 0.1 M ammonium acetate in cold methanol, and two with cold 80% acetone, and centrifuged at 14,000×
g for 7 min. The final pellet containing purified protein was dried and dissolved in the Laemmli 1DE separation buffer overnight [
30]. Proteins were then quantified by measuring the absorbance at 595 nm according to the Bradford assay [
31]. Protein yield was calculated as the mg of protein for g fresh tissue weight of each biological replicate.
4.4.2. Protein Extraction and Purification from Roots
Proteins from the roots of three biological replicates of the control and FCA treatments were extracted by 1 g of root tissue, weighed, and then pulverized in liquid nitrogen. A total of 1 mL of extraction buffer (0.7 M sucrose, 0.5 M Tris, 30 mM HCl, 50 mM EDTA, 0.1 M KCl, 2% 2-mercaptoethanol, 2 mM PMSF) was added to the pulverized tissue. The phenolic phase was performed by adding to the sample 500 µL of SDS Buffer (30% Sucrose, 2% SDS to be dissolved in 0.1 M Tris-HCl pH 8, 5% of 2-mercaptoethanol) and 500 µL of Phenol solution Sigma-Aldrich (equilibrated with 10 mM Tris HCl pH8 and 1 mM EDTA) [
32]. The sample was shaken and centrifuged at 13,000 rpm for 8 min. The supernatant phenolic phase was recovered to which 0.1 M of ammonium acetate in cold methanol was added. The sample was placed at −20 °C for 30 min, then centrifuged at 13,000 rpm for 5 min. A second wash in ammonium acetate was performed, followed by two washes with 80% acetone. The final pellet was dried and then dissolved in the Laemmli 1DE separation buffer overnight [
30]. Proteins were then quantified by measuring the absorbance at 595 nm according to the Bradford assay [
31]. Protein yield was calculated as the mg of protein for g fresh tissue weight of each biological replicate.
4.5. SDS-PAGE Electrophoresis of Proteins in Gel Digestion, and Mass Spectrometry
A gel was prepared at a concentration of 12.5% in the running gel and 6% in the stacking gel of acrylamide/bisacrylamide, according to the method of Laemmli [
30]. The samples were heated for 4 min at 100 °C before being loaded onto the gel. The electrophoretic run was conducted at 60 mA for the stacking gel and 120 mA in the running gel at a constant power of 200 V.
The electrophoresis ran for an average of 1 h and 30 min. The gels were stained with Coomassie Blue overnight and were subsequently destained with several changes of destaining solution (45% methanol, 10% acetic acid). Digitalized images of the SDS-PAGEs were analyzed by the Quantity One 1-D Analysis Software (Bio-Rad, Hercules, CA, USA) to measure the optical densities at each lane of all biological replicates. Each lane of the same SDS-PAGE was divided into six slices from 200 to10 kDa and were subsequently manually excised from the gel.
The CBB-stained gel slices were destained and then processed with the reduction (DTT) and alkylation (IAA) steps [
33]. Gel pieces were digested by trypsin (Promega, Madison, WI, USA) overnight at 37 °C, followed by the addition of an ammonium bicarbonate buffer to cover the gel matrix. The extracted peptides were immediately processed for mass spectrometry analysis.
4.6. Mass Spectrometric Analysis
LC-MS/MS analysis was performed on an EASY-LC 1000 (Thermo Fisher Scientific, Odensem, Denmark) coupled to a hybrid quadrupole/Orbitrap Q-Exactive mass spectrometer (Thermo Fisher Scientific, Dreieich, Germany). An in-house made analytical column (length 14 cm, and inner diameter 75 μm) packed with 3 μm C 18 silica particles (Dr. Maisch, Entringen, Germany) was used. Samples were diluted 5-fold in 0.1% formic acid; then, 2 μL of the resulting peptide mix was injected for LC-MS/MS analysis. Mobile phase A was 2% acetonitrile, 0.1% formic acid; mobile phase B was 80% acetonitrile, 0.1% formic acid. The LC mobile phase composition went from 0% mobile phase B to 3% mobile phase B in 1 s, then from 3 to 40% B in 120 min, and then to 100% B following an additional 8 min; after 5 min at 100% B, the mobile phase composition was brought back to 0% B in 2 min, for a total run time of 135 min at a flow rate of 230 nL/min. The column effluent was subjected to nano-electrospray ionization (1600 V of nESI potential), and the resulting charged species were detected by the Q-Exactive hybrid mass spectrometer operating in the positive ion mode. A full MS scan was acquired in the Orbitrap analyzer at a resolution of 70,000, m/z range of 350–1800, and target AGC value of 1.00 × 10−6, respectively, The data-dependent MS/MS acquisition (DDA) procedure was performed by selecting the 12 most abundant peaks with more than two charges after each full scan analysis (top 12 method). Precursor ions were fragmented by HCD (high-energy collisional dissociation); HCD normalized collision energy was calculated as 25%. MS/MS analysis was conducted in the Orbitrap analyzer at a resolution of 35,000,target AGC value of 1.0 × 10−5, and an intensity threshold of 5.0 × 10−4; the isolation window was set to 1.6 m/z. A maximum injection time of 50 ms was set for the full MS scan event, while 120 ms was the maximum injection time allowed for tandem MS/MS scans. Dynamic exclusion time was set to 30 s.
4.7. Bioinformatic Analysis and Proteins Identification
From the MS/MS spectra, protein inference and validation were performed with the Scaffold software 4.8 (Proteome Software, Inc., Portland, OR, USA). MS/MS spectra were extracted from the raw data by accepting one minimum sequence of eight amino acids, and fusion scans with the same precursor within one mass window of ±0.4 m/z over a time interval of ±30 s. The key parameters of research were the scored peak intensity, (SPI) ≥ 50%, the precursor mass tolerance of ±10 ppm, and the mass tolerance of product ions of ±20 ppm. The carbamidomethylating of cysteine was fixed as a modification, and trypsin was selected as the enzyme for the digestion, accepting two missing cleavages per peptide.
The automatic thresholds were used for peptide identification in the software Scaffold. Generally, peptide probabilities are evaluated using a Bayesian approach for the estimation of the local FDR (LFDR) up to a value of 1%. The peptide sequences using the Scaffold 4.8 Q+S system software were interfaced with both the database of proteins deduced from the generalist protein sequences of
Lactuca sativa deposited in the NCBI database (downloaded in June 2022) and in the bank UniProt data (downloaded in June 2022). Identified peptides assigned to each protein in all samples and the related statistical parameters for significant identification are reported in
Supplementary Tables S2 and S3.
4.8. Semi-Quantitative Analysis of Identified Proteins
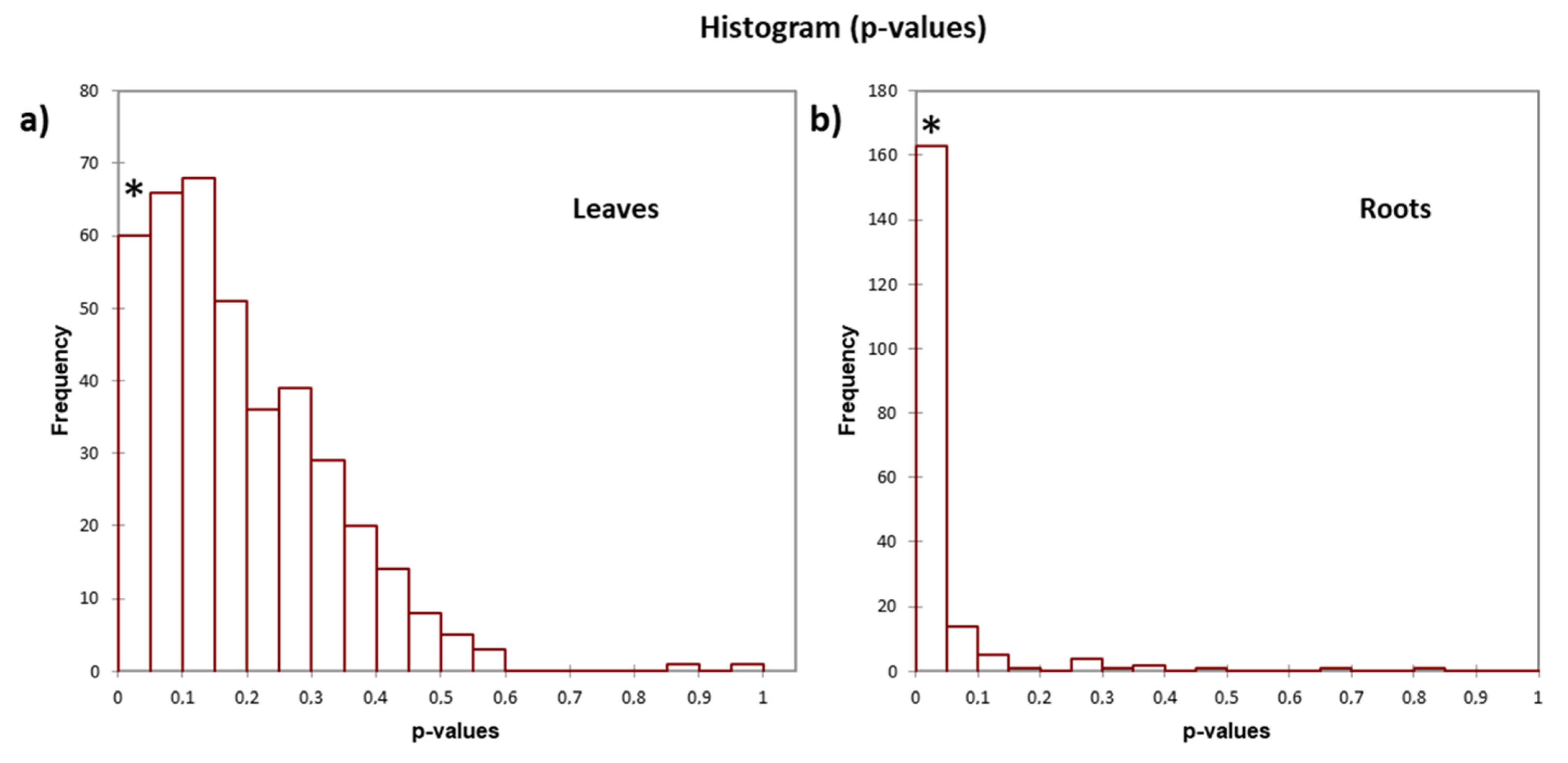
Three biological replicates of leaves and three of roots for each test were used for quantitative analyzes. The relative abundance of proteins among the samples was performed by choosing the label free quantitative method “Total Spectra” from the Quantitative menu of the Scaffold software (Proteome Software, Inc., Portland, OR, USA; version 5.1). This method uses the sum of all weighted spectra that are associated with a specific protein and within a sample, where the weight is a measure of how much a spectrum is shared by other proteins. Spectral count was undertaken only on statistically validated spectra to increase its accuracy. Consequently, it was used for quantitation comparisons. A peptide with less than two matches was discarded. The missing values were considered undetectable and were thus assumed that they were under the limit of detection, but still present. Thus, when they were undetectable, a zero value was attributed, and they were considered in the statistical calculation. To identify the proteins which show different quantitative abundances in two or more categories, the test “Fold change by category” was used. Fold change (FC) is expressed as the ratio of the quantitative value in one sample (or category) over the quantitative value in a second sample. Values of FC > 1 indicate a high quantitative profile and protein results as accumulated, while values of FC < 1 indicate a low quantitative profile and protein results as depleted. As the specified minimum value replaces any missing values, if a zero appears in the denominator an INF will appear in the FC column. The FC has also been log2 normalized for this study, and analysis was conducted using a threshold of 2 for the significance of the quantitative profile.
4.9. Gene Ontology Categories and PANTHER and KEGG Pathway Enrichment Analysis
Gene ontology (GO) categories of all differential accumulated proteins were assigned by means of the panther classification system [
34]; then, a statistical assessment of the differences in functional classes between two groups of sequences based on the Fisher test analysis were executed against the complete dataset of sequences of
Lactuca sativa. Three different significance parameters are given for the false-positive control: false discovery rate (FDR), family-wise error rate (FWER), and a single test
p-value (Fisher
p-value). By taking an FDR significance threshold of 0.05, we obtain functionalities that are specific for the organs and significant for the proteins in the treated leaves and roots. The relative fold-change of each GO term has been represented for the “Biological process” categories.
The list of FASTA sequences of DAPs whose variation patterns were highly significant (
p < 0.05) were subjected to BlastKOALA (
http://www.kegg.jp/blastkoala) (accessed on 27 May 2023) [
35] analysis to obtain KEGG Orthology (KO) assignments. The KO identifier (called the K number) list was then used for KEGG pathway mapping through the KEGG mapper web server (
http://www.genome.jp/kegg/tool/map_pathway2.html) (accessed on 27 May 2023) [
36].
4.10. Statistics
Comparison of the differences among the groups of values for the biomass and photosynthetic pigment were analyzed using the t-test with a p < 0.05 threshold for statistical significance. All the statistical analyzes were performed using Excel XLSTAT (©Addinsoft, Paris, France, released at 2022.6.1.1187). Significance was defined as p ≤ 0.05.
For the proteomics results, comparison of differences among the groups was conducted using the differentially expression and heat map tools available at XLSTAT. The Bonferroni test was used to test for the assumption of homogeneity of variances. Threshold for significance was p ≤ 0.05.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







