
Root and Shoot Growth of a Modern and an Old Tall Durum Wheat (Triticum durum Desf.) Variety under Dual-Purpose Management

 and
and 
Abstract
1. Introduction
2. Results
2.1. Fodder, Grain Yield and Weather
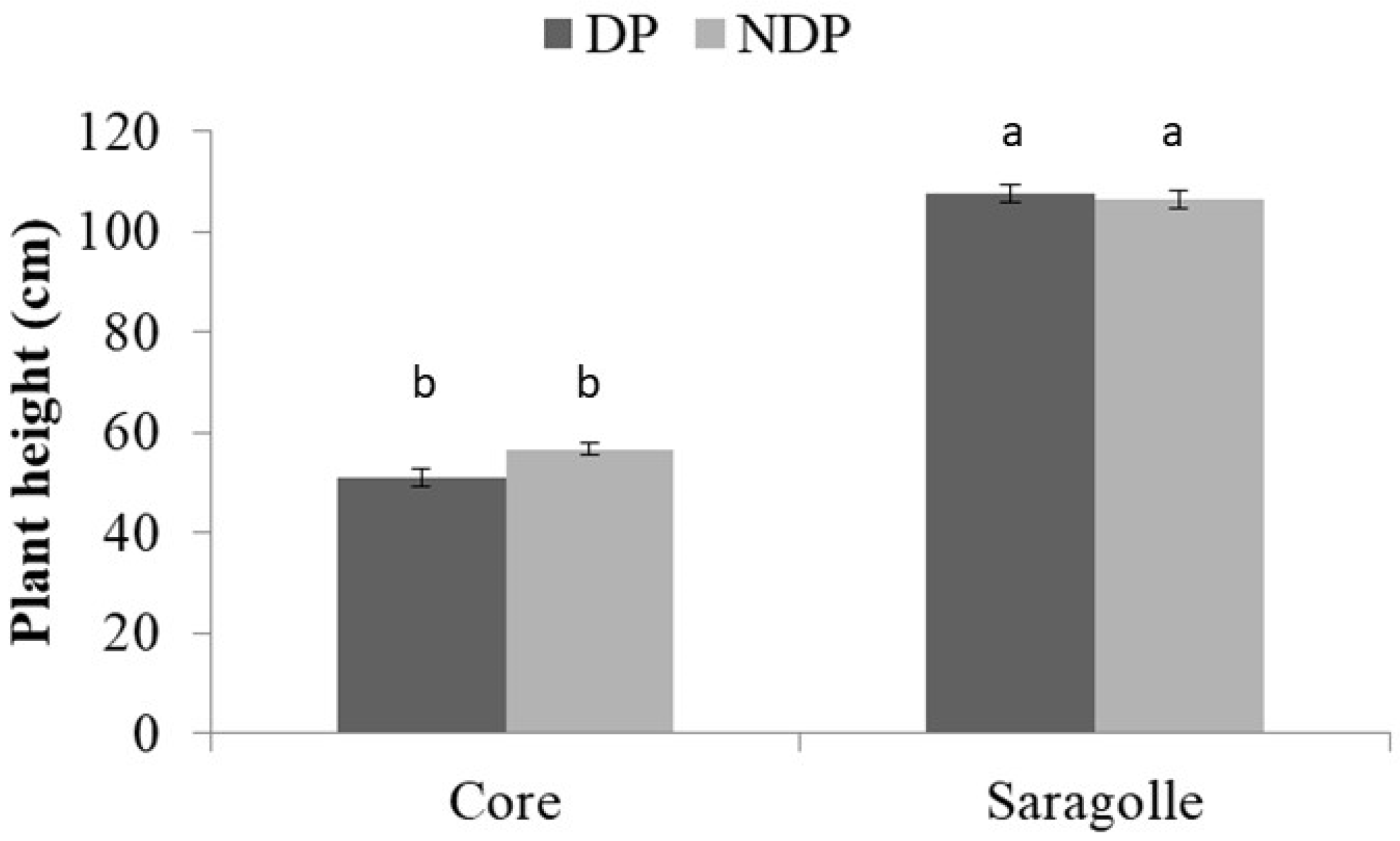
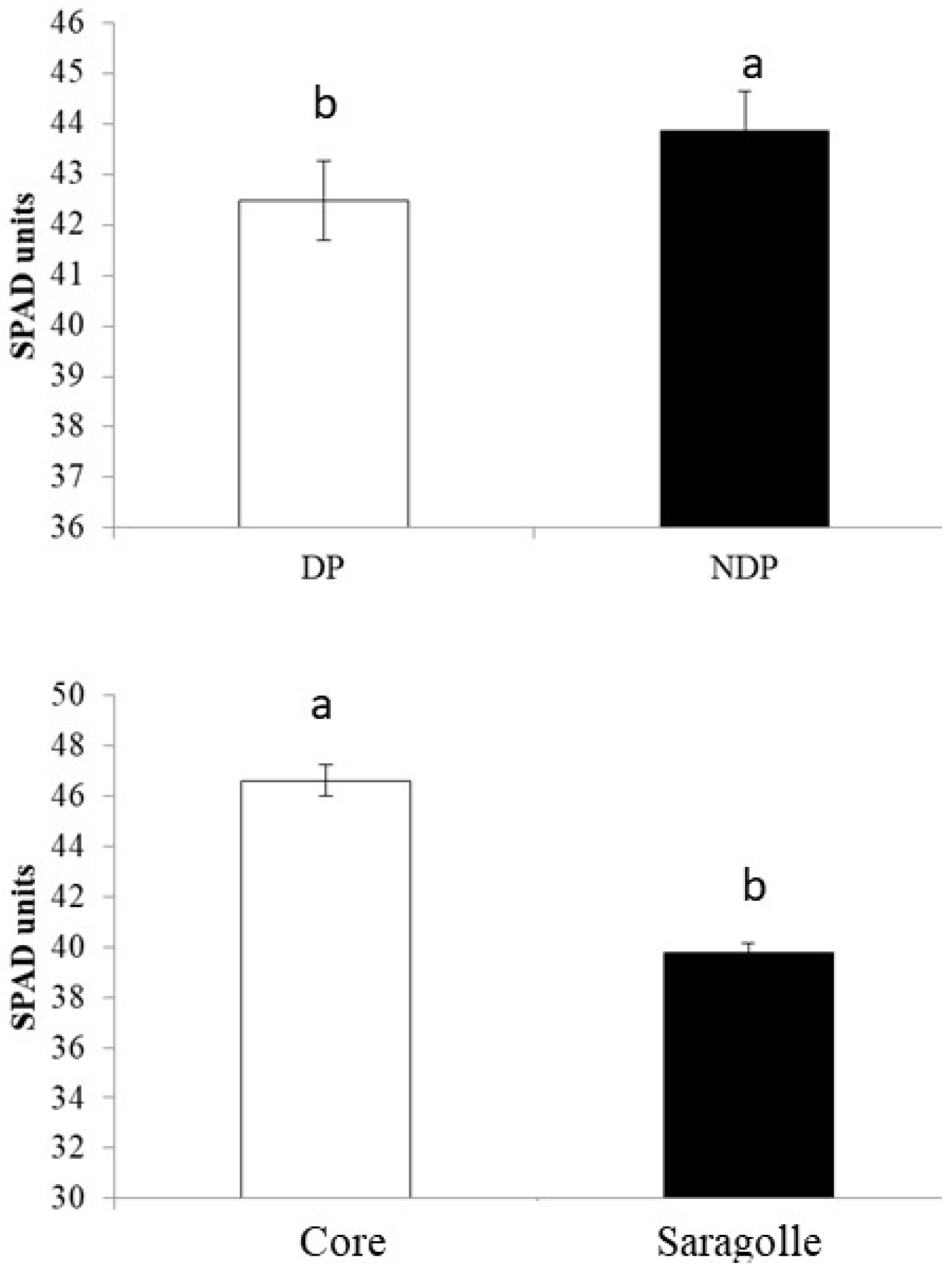
2.2. Phenology and SPAD Index
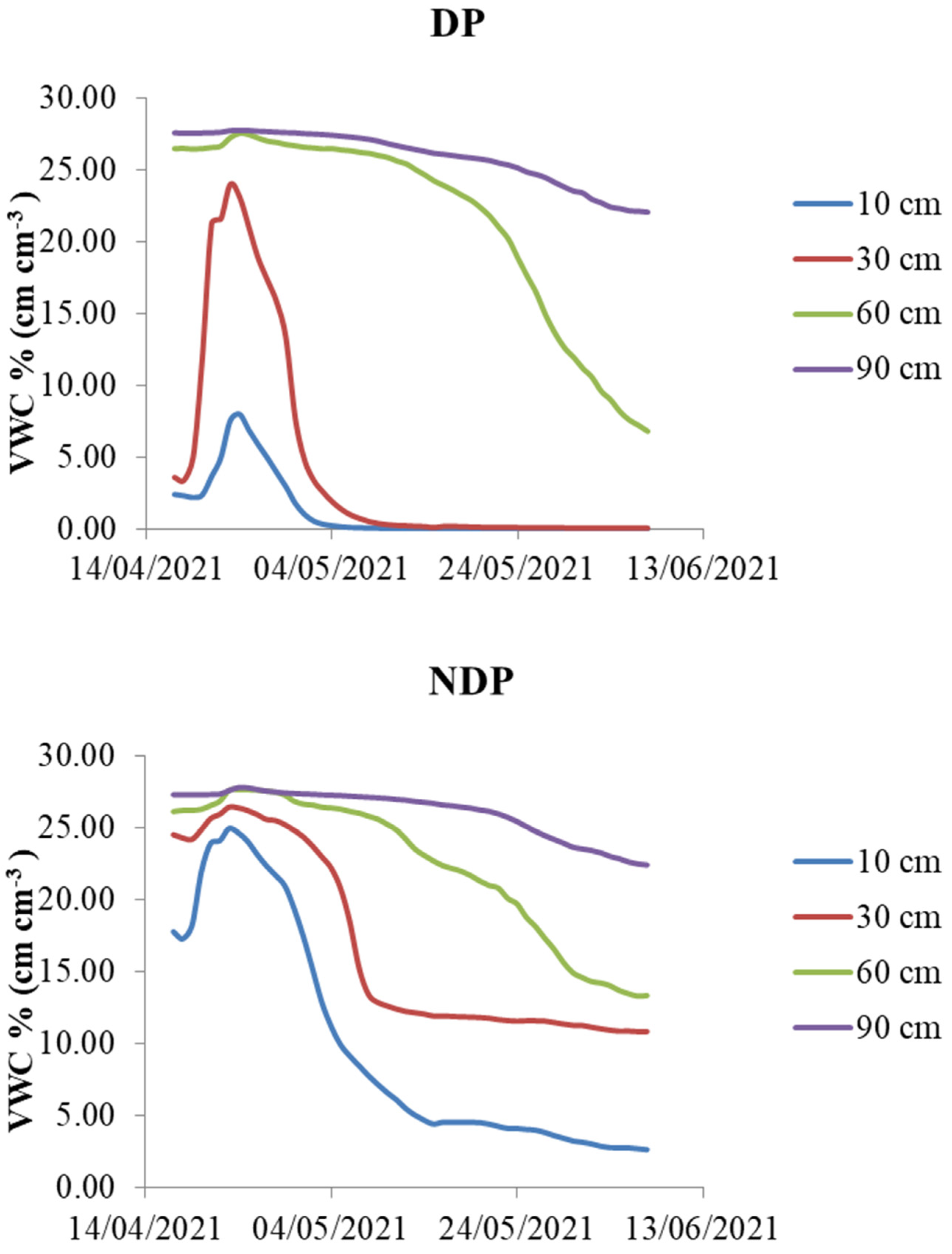
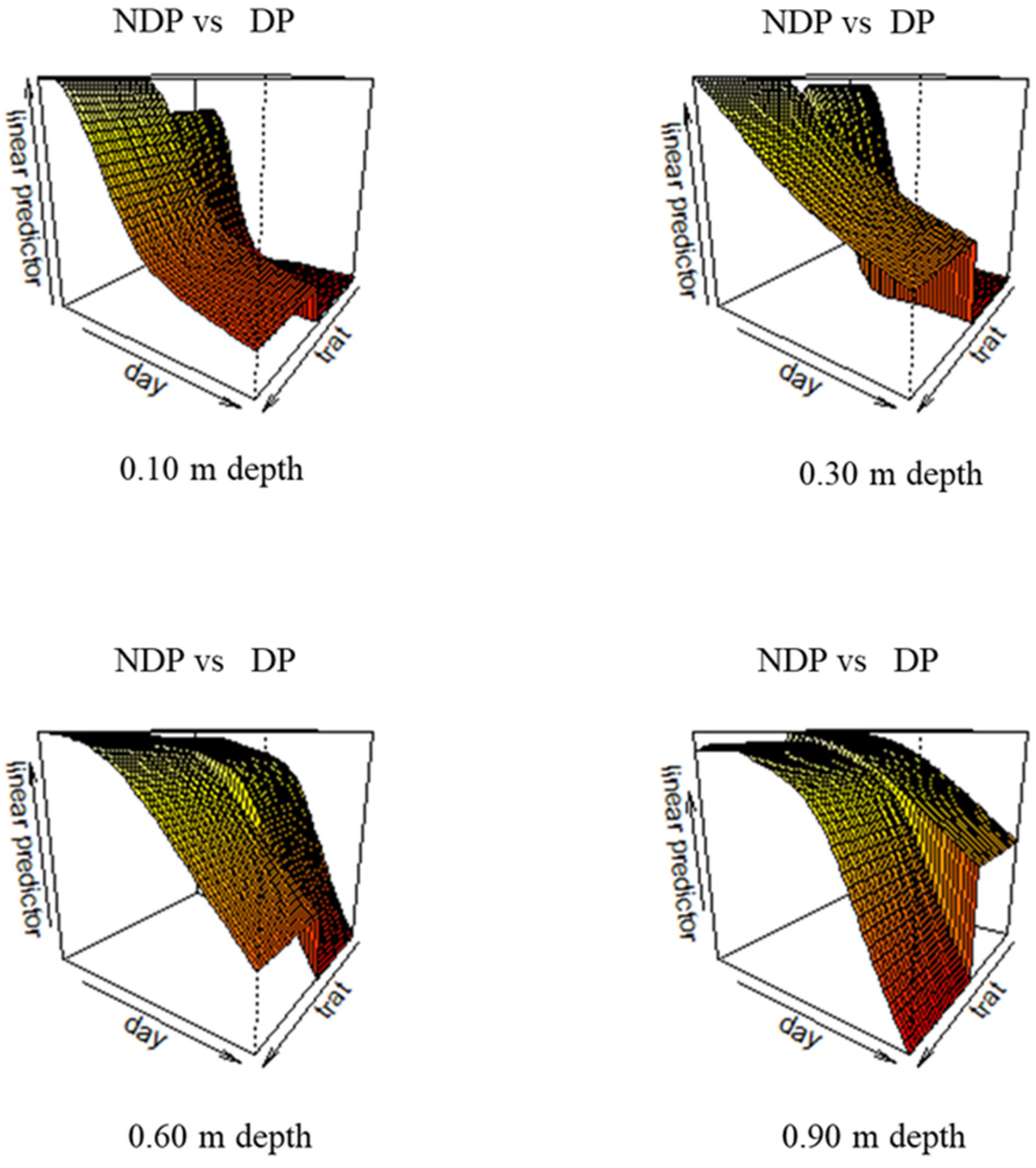
2.3. Stomatal Conductance and Soil Moisture during the Reproductive Stage
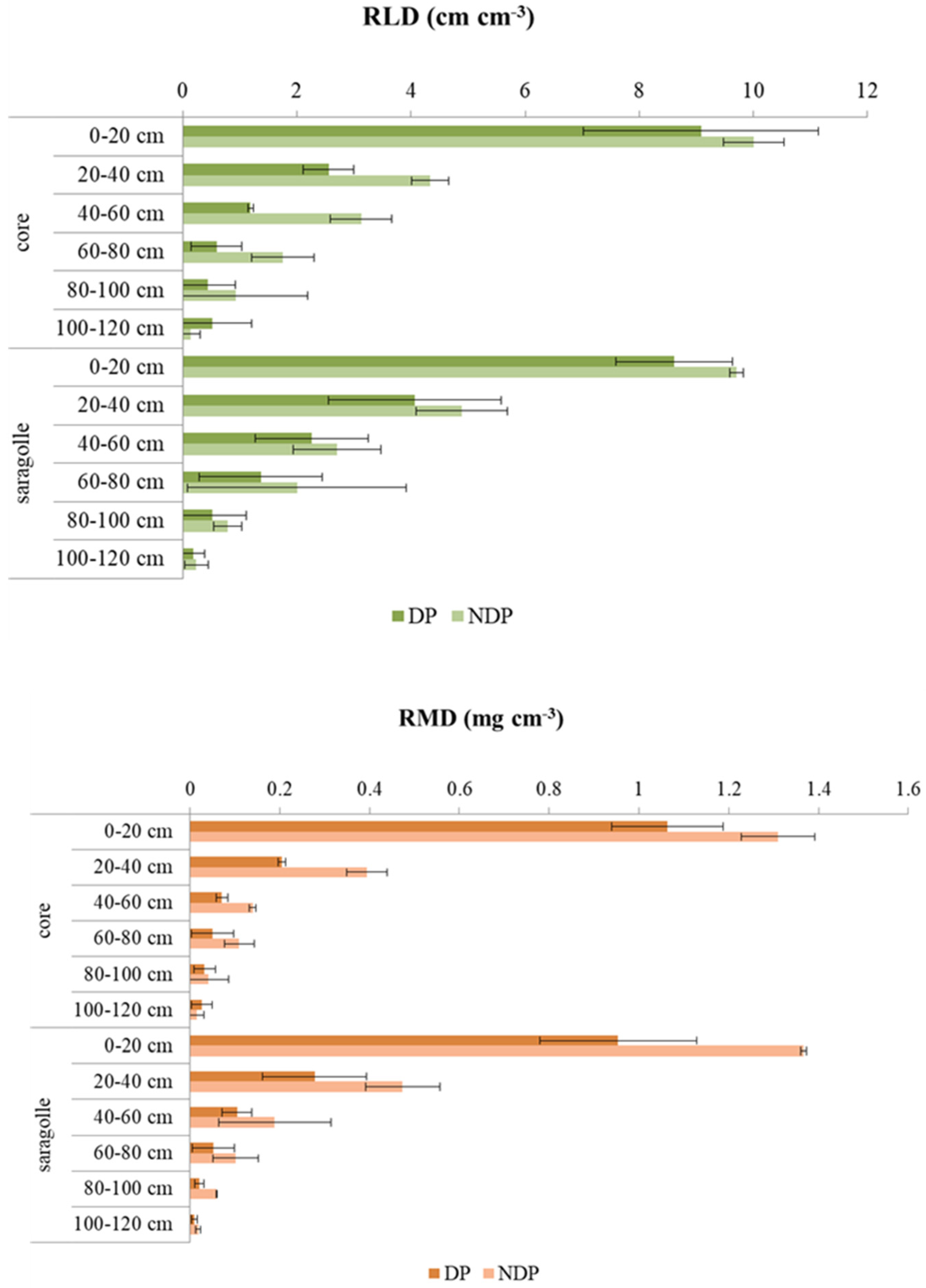
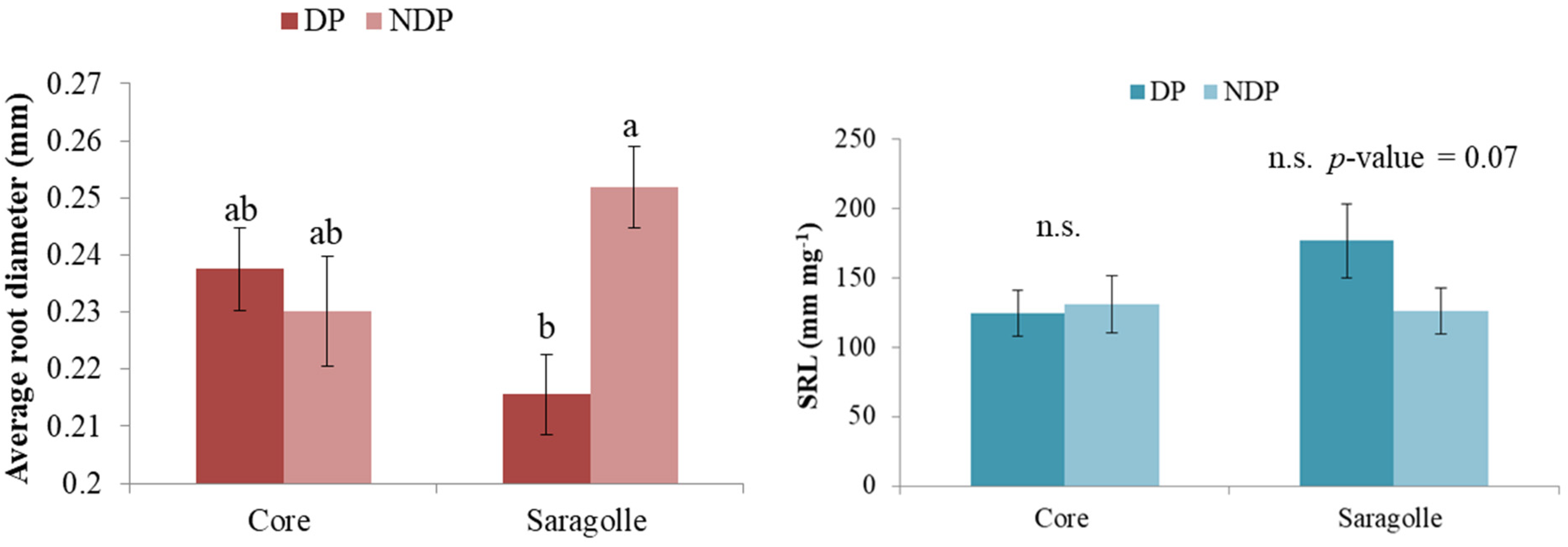
2.4. Root Biometry
3. Discussion
3.1. Fodder Biomass, Grain Yield and Weather Pattern
3.2. Phenology and SPAD Index
3.3. Stomatal Conductance and Soil Water during the Reproductive Stage
3.4. Root Biometry
4. Conclusions
- -
- One early defoliation event did not penalize grain yield. Both modern and ancient wheat fully recovered canopy losses. Defoliation delayed phenology but had no consequences on the final number of spikes per square meter.
- -
- The two varieties showed differences in leaf chlorophyll content, which was marginally affected by clipping. Stomatal conductivity did not differ between varieties; both maintained high transpiration rates throughout the reproductive season.
- -
- Our partial data (only available for modern wheat) show that soil water depletion was influenced by clipping. Defoliation did not result in a reduction in pre-anthesis water reserves, possibly due to a compensation of lower plant water demand with higher evaporative losses and delayed phenology.
- -
- Dual-purpose management markedly affected the root sync, and root mass was severely reduced by clipping. Defoliation also reduced root length, but this was more drastic for modern wheat. Saragolle lucana variety when defoliated shifted to thinner roots so root length was less impacted than root mass: a mechanism that can contribute to reducing the metabolic cost of root exploration without compromising root foraging capacity.
5. Materials and Method
5.1. Experimental Setup
5.2. Fodder Biomass, Grain Yield and Weather Pattern
5.3. Phenology and SPAD Index
5.4. Stomatal Conductance and Soil Water during the Reproductive Stage
5.5. Root Biometry
- -
- Root length density (cm cm−3) (RLD);
- -
- Root volume (cm3 cm−3);
- -
- Root average diameter (mm);
- -
- Root mass density (g cm−3) (RMD);
- -
- Specific root length (mm mg−1) (SRL).
5.6. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Aknowledgment
Conflicts of Interest
References
- Caballero, R. Castile—La Mancha: A once traditional and integrated cereal—Sheep farming system under change. Am. J. Altern. Agric. 1999, 14, 188–192. [Google Scholar] [CrossRef]
- Dove, H.; Kirkegaard, J.A. Using dual-purpose crops in sheep-grazing systems: A review. J. Sci. Food Agric. 2014, 94, 1276–1283. [Google Scholar] [CrossRef]
- Bell, L.W.; Harrison, M.T.; Kirkegaard, J.A. Dual-purpose cropping—Capitalising on potential grain crop grazing to enhance mixed-farming profitability. Crop Pasture Sci. 2015, 66, i–iv. [Google Scholar] [CrossRef]
- Kingwell, R.; Squibb, L. The role and value of combining dual-purpose crops and lucerne in a mixed-enterprise farming system. Crop Pasture Sci. 2015, 66, 399–409. [Google Scholar] [CrossRef]
- McGrath, S.R.; Pinares-Patiño, C.S.; McDonald, S.E.; Kirkegaard, J.A.; Simpson, R.J.; Moore, A.D. Utilising dual-purpose crops in an Australian high-rainfall livestock production system to increase meat and wool production. 1. Forage production and crop yields. Anim. Prod. Sci. 2021, 61, 1062–1073. [Google Scholar] [CrossRef]
- Moore, A.D.; Bell, L.W.; Revell, D.K. Feed gaps in mixed-farming systems: Insights from the Grain & Graze program. Anim. Prod. Sci. 2009, 49, 736–748. [Google Scholar]
- Dove, H.; Holst, P.J.; Stanley, D.F.; Flint, P.W. Grazing value of dual purpose wheats for young sheep. Anim. Prod. Aust. 2002, 24, 53–56. [Google Scholar]
- Dove, H.; Kirkegaard, J.A.; Kelman, W.M.; Sprague, S.J.; McDonald, S.E.; Graham, J.M. Integrating dual-purpose wheat and canola into high-rainfall livestock systems in south-eastern Australia. 2. Pasture and livestock production. Crop Pasture Sci. 2015, 66, 377–389. [Google Scholar] [CrossRef]
- Bell, L.W.; Moore, A.D.; Thomas, D.T. Diversified crop-livestock farms are risk efficient in the face of price and production variability. Agric. Syst. 2021, 87, 103050. [Google Scholar] [CrossRef]
- Butchee, D.J.; Edwards, J.T. Dual-Purpose Wheat Grain Yield as Affected by Growth Habit and Simulated Grazing Intensity. Crop Sci. 2013, 53, 1686–1692. [Google Scholar] [CrossRef]
- Kelman, W.M.; Dove, H. Growth and phenology of winter wheat and oats in a dual-purpose management system. Crop Pasture Sci. 2009, 60, 921–932. [Google Scholar] [CrossRef]
- Munsif, F.; Arif, M.; Jan, M.T.; Khan, M.J. Phenology of dual purpose wheat cultivars as influenced by planting date. Sch. J. Agric. Sci. 2013, 3, 340–350. [Google Scholar]
- Munsif, F.; Arif, M.; Jan, M.T.; Kwasar, A.; Khan, M.J. Influence of sowing dates on phenological development and yield of dual purpose wheat cultivars. Pak. J. Bot. 2015, 47, 83–88. [Google Scholar]
- Yang, J.; Lai, X.; Shen, Y. Response of dual-purpose winter wheat yield and its components to sowing date and cutting timing in a semiarid region of China. Crop Sci. 2022, 62, 425–440. [Google Scholar] [CrossRef]
- Carver, B.F.; Khalil, I.; Krenzer, E.G.; MacKown, C.T. Breeding winter wheat for a dual-purpose management system. Euphytica 2001, 119, 231–234. [Google Scholar] [CrossRef]
- Giunta, F.; Cabigliera, A.; Virdis, A.; Motzo, R. Dual-purpose use affects phenology of triticale. Field Crops Res. 2015, 183, 111–116. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Lilley, J.M.; Howe, G.N.; Graham, J.M. Impact of subsoil water use on wheat yield. Aust. J. Agric. Res. 2007, 58, 303–315. [Google Scholar] [CrossRef]
- Virgona, J.M.; Gummer, F.A.J.; Angus, J.F. Effects of grazing on wheat growth, yield, development, water use, and nitrogen use. Aust. J. Agric. Res. 2006, 57, 1307–1319. [Google Scholar] [CrossRef]
- Bonachela, S.; Orgaz, F.; Fereres, E. Winter cereals grown for grain and for the dual purpose of forage plus grain II. Water use and water-use efficiency. Field Crops Res. 1995, 44, 13–24. [Google Scholar] [CrossRef]
- Schuster, J.L. Root development of native plants under three grazing intensities. Ecology 1964, 45, 63–70. [Google Scholar] [CrossRef]
- Hodgkinson, K.C.; Becking, H.G.B. Effect of defoliation on root growth of some arid zone perennial plants. Aust. J. Agric. Res. 1978, 29, 31–42. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Liao, F.; Blancaflor, E.B. Two Wheat Cultivars with Contrasting Post-Embryonic Root Biomass Differ in Shoot Re-Growth after Defoliation: Implications for Breeding Grazing Resilient Forages. Plants 2019, 8, 470. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Lilley, J.M.; Hunt, J.R.; Sprague, S.J.; Ytting, N.K.; Rasmussen, I.S.; Graham, J.M. Effect of defoliation by grazing or shoot removal on the root growth of field-grown wheat (Triticum aestivum L.). Crop Pasture Sci. 2015, 66, 249–259. [Google Scholar] [CrossRef]
- Narayanan, S.; Mohan, A.; Gill, K.S.; Prasad, P.V.V. Variability of Root Traits in Spring Wheat Germplasm. PLoS ONE 2014, 9, e100317. [Google Scholar] [CrossRef] [PubMed]
- Wasson, A.P.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Richards, R.A.; Watt, M. Soil coring at multiple field environments can directly quantify variation in deep root traits to select wheat genotypes for breeding. J. Exp. Bot. 2014, 65, 6231–6249. [Google Scholar] [CrossRef] [PubMed]
- Francesco, G.; Pruneddu, G.; Cadeddu, F.; Motzo, R. Old tall durum wheat cultivars are suited for dual-purpose utilization. Eur. J. Agron. 2017, 5, 67–77. [Google Scholar]
- Cadeddu, F.; Motzo, R.; Mureddu, F.; Giunta, F. Ancient wheat species are suitable to grain-only and grain plus herbage utilisations in marginal Mediterranean environments. Agron. Sustain. Dev. 2021, 41, 15. [Google Scholar] [CrossRef]
- Bektas, H.; Hohn, C.E.; Waines, J.G. Root and shoot traits of bread wheat (Triticum aestivum L.) landraces and cultivars. Euphytica 2016, 212, 297–311. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Sprague, S.J.; Kirkegaard, J.A.; Bell, L.W.; Seymour, M.; Ryan, J.G.M. Dual-purpose cereals offer increased productivity across diverse regions of Australia’s high rainfall zone. Field Crops Res. 2018, 227, 119–131. [Google Scholar] [CrossRef]
- Francia, E.; Pecchioni, N.; Nicosia, O.L.D.; Paoletta, G.; Taibi, L.; Franco, V.; Odoardi, M.; Stanca, A.; Delogu, G. Dual-purpose barley and oat in a Mediterranean environment. Field Crops Res. 2006, 99, 158–166. [Google Scholar] [CrossRef]
- Harrison, M.T.; Evans, J.R.; Dove, H.; Moore, A.D. Recovery dynamics of rainfed winter wheat after livestock grazing 1. Growth rates, grain yields, soil water use and water-use efficiency. Crop Pasture Sci. 2011, 62, 947–959. [Google Scholar] [CrossRef]
- Bell, L.W.; Lilley, J.M.; Hunt, J.R.; Kirkegaard, J.A. Optimising grain yield and grazing potential of crops across Australia’s high-rainfall zone: A simulation analysis. 1. Wheat. Crop Pasture Sci. 2015, 66, 332–348. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Motzo, R. Grain yield and grain protein of old and modern durum wheat cultivars grown under different cropping systems. Field Crops Res. 2019, 230, 107–120. [Google Scholar] [CrossRef]
- De Vita, P.; Nicosia, O.L.D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding progress in morpho-physiological, agronomical and qualitative traits of durum wheat cultivars released in Italy during the 20th century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Tian, L.H.; Bell, L.W.; Shen, Y.Y.; Whish, J.P.M. Dual-purpose use of winter wheat in western China: Cutting time and nitrogen application effects on phenology, forage production, and grain yield. Crop Pasture Sci. 2012, 63, 520–528. [Google Scholar] [CrossRef]
- Edwards, J.T.; Carver, B.F.; Horn, G.W.; Payton, M.E. Impact of Dual-Purpose Management on Wheat Grain Yield. Crop Sci. 2011, 51, 2181–2185. [Google Scholar] [CrossRef]
- Drake, D.J.; Orloff, S.B. Simulated grazing effects on triticale forage yield. Forage Grazinglands 2005, 3, 1–6. [Google Scholar] [CrossRef]
- Yashawanthkumar, K.J.; Vijendra, S.B.; Sudhir, N.; Ravindra, M.P.; Juned, H.B.; Deepak, N.B.; Vittal, D.G.; Gopalareddy, K.; Chandra, N.M.; Marutha, H.M.; et al. Impact of heat and drought stress on phenological development and yield in bread wheat. Plant Phys. Rep. 2021, 26, 357–367. [Google Scholar] [CrossRef]
- Munsif, F.; Arif, M.; Khan, A.; Akhtar, K.; Ali, K.; Vincevica-Gaile, Z.; Rasul, F.; Iqbal, A.; Fahad, S.; Burlakovs, J.; et al. Dual-purpose wheat technology: A tool for ensuring food security and livestock sustainability in cereal-based cropping pattern. Arch. Agron. Soil Sci. 2021, 67, 1889–1900. [Google Scholar] [CrossRef]
- Li, J.; Lu, X.; Ju, W.; Li, J.; Zhu, S.; Zhou, Y. Seasonal changes of leaf chlorophyll content as a proxy of photosynthetic capacity in winter wheat and paddy rice. Ecol. Ind. 2022, 140, 109018. [Google Scholar] [CrossRef]
- Carucci, F.; Gatta, G.; Gagliardi, A.; De Vita, P.; Bregaglio, S.; Giuliani, M.M. Agronomic Strategies to Improve N Efficiency Indices in Organic Durum Wheat Grown in Mediterranean Area. Plants 2021, 10, 2444. [Google Scholar] [CrossRef] [PubMed]
- Bochicchio, R.; Labella, R.; Vitti, A.; Nuzzaci, M.; Logozzo, G.; Amato, M. Root Morphology, Allometric Relations and Rhizosheath of Ancient and Modern Tetraploid Wheats (Triticum durum Desf.) in Response to Inoculation with Trichoderma harzianum T-22. Plants 2022, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Ruisi, P.; Ingraffia, R.; Urso, V.; Giambalvo, D.; Alfonzo, A.; Corona, O.; Settanni, L.; Frenda, A.S. Influence of grain quality, semolinas and baker’s yeast on bread made from old landraces and modern genotypes of Sicilian durum wheat. Food Res. Int. 2021, 140, 110029. [Google Scholar] [CrossRef]
- LeCain, D.R.; Morgan, J.A.; Zerbi, G. Leaf anatomy and gas exchange in nearly isogenic semidwarf and tall winter wheat. Crop Sci. 1989, 29, 1246–1251. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, A.L. Wheat Yield Progress Associated with Higher Stomatal Conductance and Photosynthetic Rate, and Cooler Canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Roche, D. Stomatal Conductance Is Essential for Higher Yield Potential of C3 Crops. Crit. Rev. Plant Sci. 2015, 34, 429–453. [Google Scholar] [CrossRef]
- Lu, Z.; Percy, R.G.; Qualset, C.O.; Zeiger, E. Stomatal Conductance Predicts Yields in Irrigated Pima Cotton and Bread Wheat Grown at High Temperatures. J. Exp. Bot. 1998, 49, 453–460. Available online: http://www.jstor.org/stable/23695978 (accessed on 1 July 2022). [CrossRef]
- Morrison, M.J.; Voldeng, H.D.; Cober, E.R. Physiological Changes from 58 Years of Genetic Improvement of Short-Season Soybean Cultivars in Canada. Agron. J. 1999, 91, 685–689. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Has long-term selection for yield in durum wheat also induced changes in leaf and canopy traits? Field Crops Res. 2008, 106, 68–76. [Google Scholar] [CrossRef]
- Nakhforoosh, A.; Bodewein, T.; Fiorani, F.; Bodner, G. Identification of water use strategies at early growth stages in durum wheat from shoot phenotyping and physiological measurements. Front. Plant Sci. 2016, 7, 1155. [Google Scholar] [CrossRef] [PubMed]
- Annicchiarico, P.; Abdelazoui, Z.; Kelkouli, M.; Zerargui, H. Grain yield, straw yield and economic value of tall and semi-dwarf durum wheat cultivars in Algeria. J. Agric. Sci. 2005, 143, 57–64. [Google Scholar] [CrossRef]
- Bonachela, S. Root growth of triticale and barley grown for grain or for forage-plus-grain in a Mediterranean climate. Plant Soil 1996, 183, 239–251. [Google Scholar] [CrossRef]
- Richards, J.H. Root growth response to defoliation in two Agropyronbunchgrasses: Field observations with an improved root periscope. Oecologia 1984, 64, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.S. Root growth of Lolium perenne L. II. Effects of defoliation and shading. N. Z. J. Agric. Res. 1971, 14, 552–562. [Google Scholar] [CrossRef]
- Evans, P.S. The effect of repeated defoliation to three different levels on root growth of five pasture species. N. Z. J. Agric. Res. 1973, 16, 31–34. [Google Scholar] [CrossRef]
- Thornton, B.; Millard, P. Effects of severity of defoliation on root functioning in grasses. Rangel. Ecol. Manag. J. Range Manag. Arch. 1996, 49, 443–447. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Passioura, J.; Carminati, A. Hydraulic processes in roots and the rhizosphere pertinent to increasing yield of water-limited grain crops: A critical review. J. Exp. Bot. 2018, 69, 3255–3265. [Google Scholar] [CrossRef]
- Schneider, H.M.; Postma, J.A.; Kochs, J.; Pflugfelder, D.; Lynch, J.P.; van Dusschoten, D. Spatio-temporal variation in water uptake in seminal and nodal root systems of barley plants grown in soil. Front. Plant Sci. 2020, 11, 1247. [Google Scholar] [CrossRef]
- Kuhlmann, H.; Barraclough, P.B. Comparison between the seminal and nodal root systems of winter wheat in their activity for N and K uptake. Z. Pflanzenernaehr. Bodenk. 1987, 150, 24–30. [Google Scholar] [CrossRef]
- Yambao, E.B.; Ingram, K.T.; Real, J.G. Root Xylem Influence on the Water Relations and Drought Resistance of Rice. J. Exp. Bot. 1992, 43, 925–932. [Google Scholar] [CrossRef]
- Bitella, G.; Rossi, R.; Bochicchio, R.; Perniola, M.; Amato, M. A Novel Low-Cost Open-Hardware Platform for Monitoring Soil Water Content and Multiple Soil-Air-Vegetation Parameters. Sensors 2014, 14, 19639–19659. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Hastie, T. Gam: Generalized Additive Models. R Package Version 1.22. 2022. Available online: https://CRAN.R-project.org/package=gam (accessed on 1 October 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield (Tons ha−1) | ||||
|---|---|---|---|---|
| DP | NDP | |||
| Variety | Mean | St.dev | Mean | St.dev |
| Core | 2.95 | 1.00 | 3.22 | 0.63 |
| Saragolla | 2.43 | 0.59 | 2.47 | 0.41 |
| Source of Variation | Pr(>F) |
|---|---|
| T °C | 0.0002 |
| Date | 0.0521 |
| Variety | 0.5476 |
| Clipping | 0.9897 |
| Date × variety | 0.9507 |
| Date × clipping | 0.4755 |
| Variety × clipping | 0.0702 |
| Date × Variety × clipping | 0.1433 |
| Parametric Coefficients | Approximate Significance of Smooth Terms: | Goodness of Fit | ||||||
|---|---|---|---|---|---|---|---|---|
| Soil Depth (m) | Parametric Term | Estimate | Pr(>|t|) | Smooth Term | edf | p-Value | R2 Adj | GCV |
| 0.1 | Intercept | 3.428 | <0.001 | s(day) DP | 5.382 | <0.0001 | 0.44 | 36.852 |
| clipping—NDP | 4.242 | <0.001 | s(day)NDP | 3.645 | <0.0001 | |||
| 0.3 | Intercept | 6.301 | <0.001 | s(day) DP | 5.607 | <0.0001 | 0.48 | 68.139 |
| clipping—NDP | 8.010 | <0.001 | s(day)NDP | 1.688 | <0.0002 | |||
| 0.6 | Intercept | 21.630 | <0.001 | s(day) DP | 5.126 | <0.0001 | 0.79 | 9.322 |
| clipping—NDP | 0.711 | <0.001 | s(day)NDP | 2.785 | <0.0001 | |||
| 0.9 | Intercept | 26.410 | <0.0001 | s(day) DP | 2.681 | <0.0001 | 0.874 | 0.55 |
| clipping—NDP | −0.933 | <0.001 | s(day)NDP | 5.204 | <0.0001 | |||
| Source of Variation | Pr(>F) | ||
|---|---|---|---|
| RLD (cm cm−3) | Depth | 1.59 × 10−8 | *** |
| Variety | 0.506854 | ||
| Clipping | 0.000376 | *** | |
| Depth × variety | 0.831928 | ||
| Depth × clipping | 0.140179 | ||
| Variety × clipping | 0.19326 | ||
| Depth × variety × clipping | 0.53003 | ||
| RMD (mg cm−3) | Depth | 2.16 × 10−12 | *** |
| Variety | 0.494564 | ||
| Clipping | 3.64 × 10−5 | *** | |
| Depth × variety | 0.710335 | ||
| Depth × clipping | 0.001257 | ** | |
| Variety × clipping | 0.308827 | ||
| Depth × variety × clipping | 0.738796 | ||
| SRL (mm mg−1) | Variety | 0.36086 | |
| Clipping | 0.1285 | ||
| Variety × clipping | 0.05413 | ||
| Average diameter (mm) | Variety | 0.99492 | |
| Clipping | 0.064932 | ||
| Variety × clipping | 0.007635 | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, R.; Bitella, G.; Bochicchio, R.; Labella, R.; Angerame, F.; Urbano, M.; Amato, M. Root and Shoot Growth of a Modern and an Old Tall Durum Wheat (Triticum durum Desf.) Variety under Dual-Purpose Management. Plants 2023, 12, 588. https://doi.org/10.3390/plants12030588
Rossi R, Bitella G, Bochicchio R, Labella R, Angerame F, Urbano M, Amato M. Root and Shoot Growth of a Modern and an Old Tall Durum Wheat (Triticum durum Desf.) Variety under Dual-Purpose Management. Plants. 2023; 12(3):588. https://doi.org/10.3390/plants12030588
Chicago/Turabian StyleRossi, Roberta, Giovanni Bitella, Rocco Bochicchio, Rosanna Labella, Francesco Angerame, Marcella Urbano, and Mariana Amato. 2023. "Root and Shoot Growth of a Modern and an Old Tall Durum Wheat (Triticum durum Desf.) Variety under Dual-Purpose Management" Plants 12, no. 3: 588. https://doi.org/10.3390/plants12030588
APA StyleRossi, R., Bitella, G., Bochicchio, R., Labella, R., Angerame, F., Urbano, M., & Amato, M. (2023). Root and Shoot Growth of a Modern and an Old Tall Durum Wheat (Triticum durum Desf.) Variety under Dual-Purpose Management. Plants, 12(3), 588. https://doi.org/10.3390/plants12030588