
Trends in Phenolic Profiles of Achillea millefolium from Different Geographical Gradients

 , ,
, ,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Phenolic Profiles of Inflorescences, Leaves, and Stems in Turkish and Lithuanian Populations
2.2. Total Phenolic Acids and Flavonoids from Two Locations
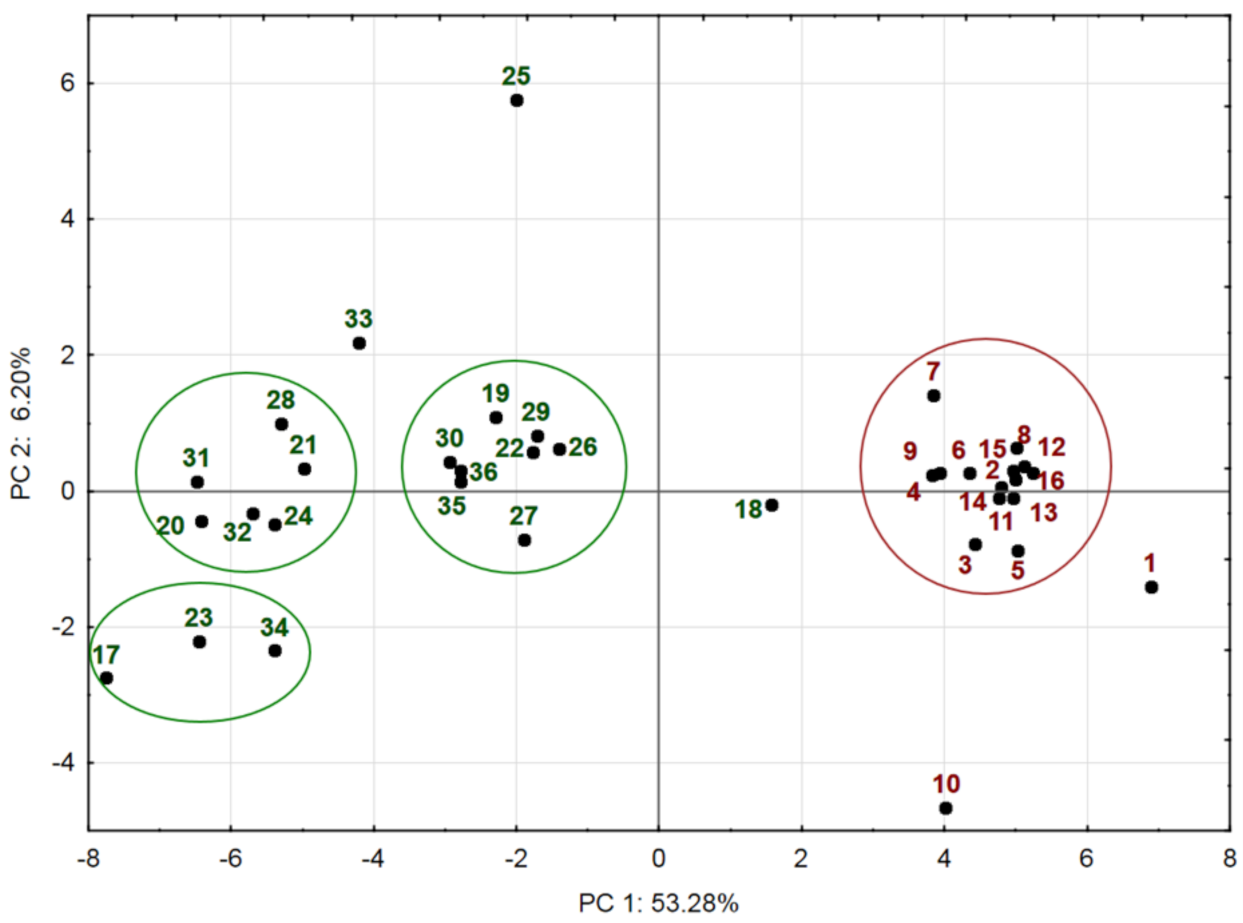
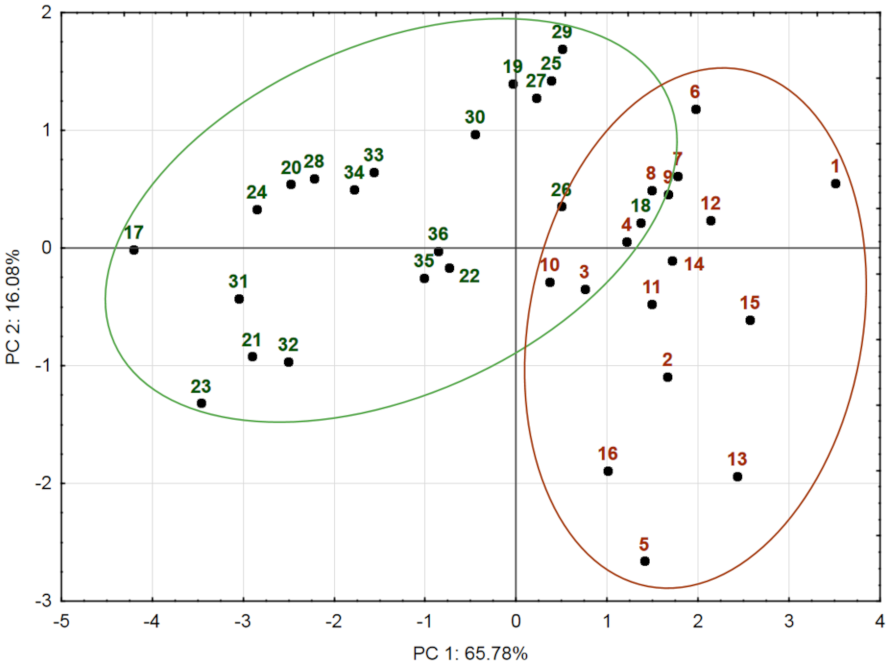
2.3. Chemical Variation of Populations: Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Plant Material, Sampling, and Identification
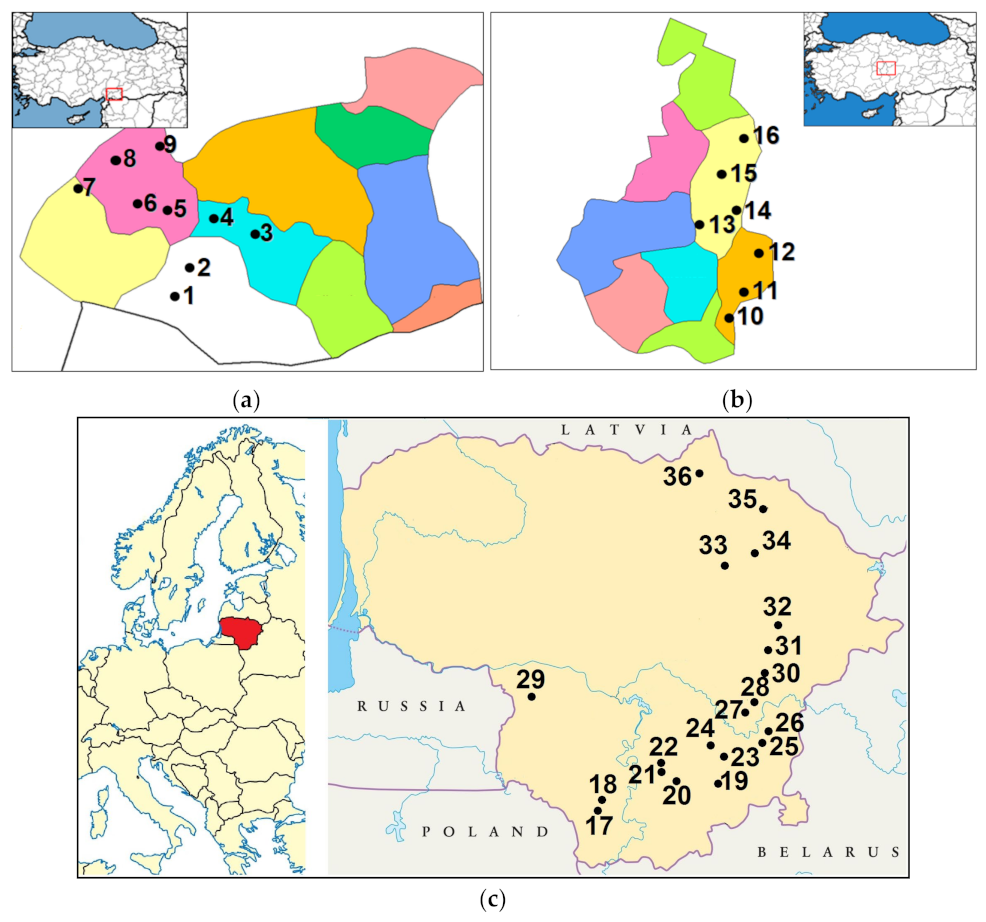
4.2. Sampling Sites
4.3. Chemicals
4.4. Extraction
4.5. Qualitative and Quantitative Analysis
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ebrahim, F.; Pakniyat, H.; Arzani, A.; Rahimmalek, M. Karyotype analysis and new chromosome number reports in Achillea species. Biologia 2012, 67, 284–288. [Google Scholar] [CrossRef]
- Richardson, I.B.K. Achillea L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: London, UK, 1976; Volume 4, pp. 159–165. [Google Scholar]
- Ehrendorfer, F.; Guo, Y.-P. Multidisciplinary studies on Achillea sensu lato (Compositae-Anthemideae): New data on systematics and phylogeography. Willdenowia 2006, 36, 69–87. [Google Scholar] [CrossRef]
- Arabacı, T. Achillea L. In Türkiye Bitkileri Listesi (Damarlı Bitkiler); Güner, A., Aslan, S., Ekim, T., Vural, M., Babaç, M.T., Eds.; Nezahat Gökyiğit Botanik Bahçesi ve Flora Araştırmaları Derneği Yayını: İstanbul, Turkey, 2012; pp. 108–112. [Google Scholar]
- Pomeroy, E.; Bennett, P.; Hunt, C.; Reynolds, T.; Farr, L.; Frouin, M.; Holman, J.; Lane, R.; French, C.; Barker, G. New Neanderthal remains associated with the ‘flower burial’ at Shanidar Cave. Antiquity 2020, 94, 11–26. [Google Scholar] [CrossRef]
- Mileffolii herba. European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/herbal/millefolii-herba (accessed on 13 December 2022).
- Mohamed, D.A.; Hanfy, E.A.; Fouda, K. Evaluation of antioxidant, anti-inflammatory and anti-arthritic activities of yarrow (Achillea millefolium). J. Biol. Sci. 2018, 18, 317–328. [Google Scholar] [CrossRef]
- Fierascu, I.; Ungureanu, C.; Avramescu, S.M.; Fierascu, R.C.; Ortan, A.; Soare, L.C.; Paunescu, A. In Vitro antioxidant and antifungal properties of Achillea millefolium L. Rom. Biotechnol. Lett. 2015, 20, 10626–10636. [Google Scholar]
- Moradi, M.T.; Rafieian-Koupaei, M.; Imani-Rastabi, R.; Nasiri, J.; Shahrani, M.; Rabiei, Z.; Alibabaeit, Z. Antispasmodic effects of yarrow (Achillea millefolium L.) extract in the isolated ileum of rat. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 499–503. [Google Scholar] [CrossRef]
- Kaczorová, D.; Karalija, E.; Dahija, S.; Bešta-Gajević, R.; Parić, A.; Zeljković, S.Ć. Influence extraction solvent on the phenolic profile and bioactivity of two Achillea species. Molecules 2021, 26, 1601. [Google Scholar] [CrossRef]
- Khan, A.U.; Gilani, A.H. Blood pressure lowering, cardiovascular inhibitory and bronchodilatory actions of Achillea millefolium. Phytother. Res. 2011, 25, 577–583. [Google Scholar] [CrossRef]
- Noureddini, M.; Rasta, V.R. Analgesic effect of aqueous extract of Achillea millefolium L. on rat’s formalin test. Pharmacologyonline 2008, 3, 659–664. [Google Scholar]
- Applequist, W.L.; Moerman, D.E. Yarrow (Achillea millefolium L.): A neglected Panacea? A review of ethnobotany, bioactivity, and biomedical research. Econ. Bot. 2011, 65, 209–225. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Dueñas, M.; Pereira, E.; Carvalho, A.M.; Alves, R.C.; Oliveira, M.B.; Santos-Buelga, C.; Ferreira, I.C. Chemical composition of wild and commercial Achillea millefolium L. and bioactivity of the methanolic extract, infusion and decoction. Food. Chem. 2013, 141, 4152–4160. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Barreira, J.C.M.; Oliveira, M.B.P.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crops Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Huyut, Z.; Beydemir, Ş.; Gülçin, I. Antioxidant and antiradical properties of selected flavonoids and phenolic compounds. Biochem. Res. Int. 2017, 2017, 6791. [Google Scholar] [CrossRef]
- Magana, A.A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Gevrenova, R.; Zengin, G.; Sinan, K.I.; Yıldıztugay, E.; Zheleva-Dimitrova, D.; Picot-Allain, C.; Mahomoodally, M.F.; Imran, M.; Dall’acqua, S. UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey. Antioxidants 2021, 10, 1180. [Google Scholar] [CrossRef]
- Sevindik, H.G.; Güvenalp, Z.; Yerdelen, K.Ö.; Yuca, H.; Demirezer, Ö. The discovery of potential anticholinesterase compounds from Achillea millefolium L. Ind. Crops Prod. 2015, 76, 873–879. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A.; Ceylan, R.; Uysal, S.; Mocan, A.; Guler, G.O.; Mahomoodally, M.F.; Glamočlija, J.; Ćirić, A.; Soković, M. Shedding light on the biological and chemical fingerprints of three Achillea species (A. biebersteinii, A. millefolium and A. teretifolia). Food Funct. 2017, 8, 1152–1165. [Google Scholar] [CrossRef]
- Afshari, M.; Rahimmalek, M.; Miroliaei, M. Variation in polyphenolic profiles, antioxidant and antimicrobial activity of different Achillea species as natural sources of antiglycative compounds. Chem. Biodivers. 2018, 15, e1800075. [Google Scholar] [CrossRef]
- Hawryl, A.; Hawrył, M.; Świeboda, R.; Waksmundzka-Hajnos, M. Chemometric analysis based on high-performance liquid chromatographic data in the fingerprint construction of selected Achillea species. J. Liq. Chromatogr. Relat. Technol. 2018, 41, 1–5. [Google Scholar] [CrossRef]
- Gaweł-Bȩben, K.; Strzȩpek-Gomółka, M.; Czop, M.; Sakipova, Z.; Głowniak, K.; Kukula-Koch, W. Achillea millefolium L. and Achillea biebersteinii Afan. hydroglycolic extracts-bioactive ingredients for cosmetic use. Molecules 2020, 25, 3368. [Google Scholar] [CrossRef]
- Georgieva, L.; Gadjalova, A.; Mihaylova, D.; Pavlov, A. Achillea millefolium L.—Phytochemical profile and in vitro antioxidant activity. Int. Food Res. J. 2015, 22, 1347–1352. [Google Scholar]
- Ahlstrand, N.I.; Reghev, N.H.; Markussen, B.; Hansen, H.C.B.; Eiriksson, F.F.; Thorsteinsdóttir, M.; Rønsted, N.; Barnes, C.J. Untargeted metabolic profiling reveals geography as the strongest predictor of metabolic phenotypes of a cosmopolitan weed. Ecol. Evol. 2018, 8, 6812–6826. [Google Scholar] [CrossRef] [PubMed]
- Grebner, D.L.; Bettinger, P.; Siry, J.P. Forest Dynamics. In Introduction to Forestry and Natural Resources; Elsevier BV: Amsterdam, The Netherlands, 2013; pp. 243–254. [Google Scholar] [CrossRef]
- Labarrere, B.; Prinzing, A.; Dorey, D.; Chesneau, E.; Hennion, F. Variations of secondary metabolites among natural populations of sub-Antarctic Ranunculus species suggest functional redundancy and versatility. Plants 2019, 8, 234. [Google Scholar] [CrossRef]
- Padilla-González, G.F.; Frey, M.; Gómez-Zeledón, J.; Da Costa, F.B.; Spring, O. Metabolomic and gene expression approaches reveal the developmental and environmental regulation of the secondary metabolism of yacón (Smallanthus sonchifolius, Asteraceae). Sci. Rep. 2019, 9, 9246. [Google Scholar] [CrossRef] [PubMed]
- Martz, F.; Peltola, R.; Fontanay, S.; Duval, R.E.; Julkunen-Tiitto, R.; Stark, S. Effect of latitude and altitude on the terpenoid and soluble phenolic composition of Juniper (Juniperus communis) needles and evaluation of their antibacterial activity in the boreal zone. J. Agr. Food Chem. 2009, 57, 9575–9584. [Google Scholar] [CrossRef] [PubMed]
- Jakaaola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef]
- Zidorn, C. Altitudinal variation of phenolics contents in flowering heads of the Asteraceae family. Phytochem. Rev. 2010, 9, 197–203. [Google Scholar] [CrossRef]
- Asensio, E.; Vitales, D.; Perez, I.; Peralba, L.; Viruel, J.; Montaner, C.; Vallès, J.; Garnatje, T.; Sales, E. Phenolic compounds content and genetic diversity at population level across the natural distribution range of bearberry (Arctostaphylos uva-ursi, Ericaceae) in the Iberian Peninsula. Plants 2020, 9, 1250. [Google Scholar] [CrossRef]
- Zou, K.; Liu, X.; Zhang, D.; Yang, Q.; Fu, S.; Meng, D.; Chang, W.; Li, R.; Yin, H.; Liang, Y. Flavonoid biosynthesis is likely more susceptible to elevation and tree age than other branch pathways involved in phenylpropanoid biosynthesis in ginkgo leaves. Front. Plant Sci. 2019, 10, 983. [Google Scholar] [CrossRef]
- Spinardi, A.; Cola, G.; Gardana, C.S.; Mignani, I. Variation of anthocyanin content and profile throughout fruit development and ripening of highbush blueberry cultivars grown at two different altitudes. Front. Plant Sci. 2019, 10, 1045. [Google Scholar] [CrossRef]
- Yuliani, Y.; Rachmadiarti, F.; Dewi, S.K.; Asri, M.T.; Soegianto, A. Total phenolic and flavonoid contents of Elephantopus scaber and Ageratum conyzoides (Asteraceae) leaves extracts from various altitude habitats. Ecol. Environ. Conserv. 2019, 25, 106–113. [Google Scholar]
- Martz, F.; Jaakola, L.; Julkunen-Tiitto, R.; Stark, S. Phenolic composition and antioxidant capacity of bilberry (Vaccinium myrtillus) leaves in Northern Europe following foliar development and along environmental gradients. J. Chem. Ecol. 2010, 36, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Filippi, A.; Braidot, E.; Petrussa, E.; Fabro, M.; Vuerich, M.; Boscutti, F. Plant growth shapes the effects of elevation on the content and variability of flavonoids in subalpine bilberry stands. Plant Biol. 2021, 23, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, N.; Hussain, M.; Ibrahim, M.; Hausmann, A.E.; Rao, S.; Kaur, S.; Khazir, J.; Mir, B.A.; Olsson, S.B. Effect of altitude on volatile organic and phenolic compounds of Artemisia brevifolia Wall ex Dc. from the Western Himalayas. Front. Ecol. Evol. 2022, 10, 864728. [Google Scholar] [CrossRef]
- Tripp, E.A.; Zhuang, Y.; Schreiber, M.; Stone, H.; Berardi, A.E. Evolutionary and ecological drivers of plant flavonoids across a large latitudinal gradient. Mol. Phylogenet. Evol. 2018, 128, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Stark, S.; Julkunen-Tiitto, R.; Holappa, E.; Mikkola, K.; Nikula, A. Concentrations of foliar quercetin in natural populations of white birch (Betula pubescens) increase with latitude. J. Chem. Ecol. 2008, 34, 1382–1391. [Google Scholar] [CrossRef]
- Lätti, A.K.; Riihinen, K.R.; Kainulainen, P.S. Analysis of anthocyanin variation in wild populations of bilberry (Vaccinium myrtillus L.) in Finland. J. Agric. Food Chem. 2008, 56, 190–196. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Külheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef]
- López-Caamal, A.; Tovar-Sánche, E. Genetic, morphological, and chemical patterns of plant hybridization. Rev. Chil. Hist. Nat. 2014, 87, 16. [Google Scholar] [CrossRef]
- Luo, J.; Wang, K.; Li, G.S.; Lei, D.Q.; Huang, Y.J.; Li, W.D.; Chen, Y.Q.; Kong, J.I. 3,5-dicaffeoylquinic acid disperses Aspergillus fumigatus biofilm and enhances fungicidal efficacy of voriconazole and amphotericin B. Med. Sci. Monit. 2018, 22, 427–437. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Cholevas, C.; Polyzoidis, K.; Politis, A. Long-COVID syndrome-associated brain fog and chemofog: Luteolin to the rescue. Biofactors 2021, 47, 232–241. [Google Scholar] [CrossRef]
- Franza, L.; Carusi, V.; Nucera, E.; Pandolfi, F. Luteolin, inflammation and cancer: Special emphasis on gut microbiota. Biofactors 2021, 47, 181–189. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Ali, S.I.; Gopalakrishnan, B.; Venkatesalu, V. Pharmacognosy, phytochemistry and pharmacological properties of Achillea millefolium L.: A Review. Phytother. Res. 2017, 31, 1140–1161. [Google Scholar] [CrossRef]
- Keser, S.; Celik, S.; Turkoglu, S.; Yilmaz, O.; Turkoglu, I. Antioxidant activity, total phenolic and flavonoid content of water and ethanol extracts from Achillea millefolium L. Turk. J. Pharm. Sci. 2013, 10, 385–392. [Google Scholar]
- Benedek, B.; Gjoncaj, N.; Saukel, J.; Kopp, B. Distribution of phenolic compounds in Middleeuropean taxa of the Achillea millefolium L. Aggregate. Chem. Biodivers. 2007, 4, 849–857. [Google Scholar] [CrossRef]
- Bączek, K.; Kosakowska, O.; Przybył, J.L.; Kuźm, P.; Ejdys, M.; Obiedzinski, M.; Weglarz, Z. Intraspecific variability of yarrow (Achillea millefolium L. s.l.) in respect of developmental and chemical traits. Herba Pol. 2015, 61, 37–52. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Zidorn, C. Guidelines for consistent characterisation and documentation of plant source materials for studies in phytochemistry and phytopharmacology. Phytochemistry 2017, 139, 56–59. [Google Scholar] [CrossRef]
- Aydın, Y. Quantification of water requirement of some major crops under semi-arid climate in Turkey. PeerJ 2022, 10, e13696. [Google Scholar] [CrossRef]
- Bağdatlı, M.C.; Arıkan, E.N. Evaluation of monthly maximum, minimum and average temperature changes observed for many years in Nevsehir province of Turkey. World Res. J. Agric. Sci. 2020, 7, 209–220. [Google Scholar]
- Raudone, L.; Radušiene, J.; Seyis, F.; Yayla, F.; Vilkickyte, G.; Marksa, M.; Ivanauskas, L.; Cırak, C. Distribution of phenolic compounds and antioxidant activity in plant parts and populations of seven underutilized wild Achillea species. Plants 2022, 11, 447. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Inflorescences (n = 36) | Leaves (n = 36) | Stems (n = 36) | F | p1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Turkey | Lithuania | p | Turkey | Lithuania | p | Turkey | Lithuania | p | |||||||||
| M | SD | M | SD | M | SD | M | SD | M | SD | M | SD | ||||||
| Neochlorogenic acid | 219.9 | 37.8 | 395.7 | 85.5 | <0.001 | 557.3 | 282.4 | 979.8 | 261.5 | <0.001 | 265.2 | 70.0 | 254.1 | 59.0 | 0.607 | 69.3 | <0.001 |
| Chlorogenic acid | 1443.8 | 736.5 | 7735.3 | 2273.2 | <0.001 | 4140.3 | 3294.7 | 19,483.3 | 6832.2 | <0.001 | 1068.6 | 610.8 | 4069.3 | 1251.3 | <0.001 | 27.6 | <0.001 |
| 4-O-caffeoylquinic acid | 1362.7 | 508.9 | 992.4 | 572.9 | 0.051 | 1536.7 | 686.6 | 408.3 | 478.9 | <0.001 | 641.5 | 237.1 | 967.4 | 791.1 | 0.122 | 2.4 | 0.097 |
| 3.4-O-dicaffeoylquinic acid | 1701.0 | 997.2 | 2582.3 | 690.5 | 0.004 | 982.9 | 869.4 | 3482.3 | 1231.2 | <0.001 | 184.8 | 116.0 | 738.1 | 378.0 | <0.001 | 30.7 | <0.001 |
| 3.5-O-dicaffeoylquinic acid | 2409.4 | 1078.6 | 8646.8 | 1750.2 | <0.001 | 3749.8 | 2156.8 | 11,901.1 | 4087.7 | <0.001 | 755.4 | 314.3 | 1448.4 | 465.3 | <0.001 | 35.4 | <0.001 |
| 1,3-O-dicaffeoylquinic acid | 0 | 0 | 0 | 0 | 0 | 0 | 82.0 | 70.3 | 0 | 0 | 37.0 | 47.3 | 14.1 | <0.001 | |||
| 1.5-O-dicaffeoylquinic acid | 0 | 0 | 2345.6 | 954.3 | <0.001 | 51.8 | 23.6 | 4924.7 | 2591.7 | <0.001 | 64.4 | 25.5 | 1163.8 | 624.4 | <0.001 | 10.2 | <0.001 |
| 4.5-O-dicaffeoylquinic acid | 318.4 | 125.5 | 1010.8 | 289.0 | <0.001 | 142.8 | 93.5 | 1820.3 | 796.7 | <0.001 | 105.0 | 62.5 | 505.2 | 284.7 | <0.001 | 11.4 | <0.001 |
| Caffeic acid | 1.6 | 6.4 | 19.6 | 13.9 | <0.001 | 35.1 | 24.02 | 70.6 | 44.6 | 0.007 | 14.5 | 17.9 | 39.5 | 29.5 | 0.005 | 19.6 | <0.001 |
| Quercitrin | 0 | 0 | 289.6 | 429.9 | 0.011 | 0 | 0 | 255.6 | 161.3 | <0.001 | 0 | 0 | 90.0 | 56.6 | <0.001 | 2.4 | 0.093 |
| Rutin | 188.5 | 120.4 | 168.7 | 180.4 | 0.709 | 1979.0 | 1340.7 | 2681.0 | 1259.8 | 0.116 | 649.4 | 369.2 | 872.6 | 511.2 | 0.152 | 69.5 | <0.001 |
| Quercetin | 34.2 | 26.1 | 26.4 | 3.6 | 0.196 | 30.1 | 14.1 | 18.6 | 2.6 | 0.001 | 30.8 | 8.8 | 22.3 | 4.8 | 0.001 | 2.1 | 0.130 |
| Isoquercitrin | 425.0 | 432.2 | 6.5 | 20.7 | <0.001 | 55.2 | 132.8 | 86.8 | 123.6 | 0.466 | 0 | 0 | 33.4 | 64.5 | 0.047 | 5.9 | 0.004 |
| Luteolin | 2699.7 | 2072.7 | 814.2 | 1126.9 | 0.001 | 395.2 | 915.7 | 146.2 | 57.2 | 0.232 | 120.3 | 36.5 | 134.4 | 9.8 | 0.107 | 20.3 | <0.001 |
| Luteolin-7-O-glucoside | 1776.1 | 954.5 | 2772.1 | 702.5 | 0.001 | 262.3 | 485.7 | 975.5 | 670.0 | 0.001 | 48.7 | 51.6 | 94.2 | 98.2 | 0.103 | 106.4 | <0.001 |
| Luteolin-7-O-rutinoside | 601.4 | 409.2 | 575.4 | 370.2 | 0.843 | 160.1 | 131.7 | 1154.4 | 511.8 | <0.001 | 214.5 | 125.2 | 195.1 | 91.3 | 0.594 | 13.6 | <0.001 |
| Luteolin-3.7-O-diglucoside | 1836.4 | 1507.8 | 0 | 0 | <0.001 | 104.6 | 418.4 | 0 | 0.0 | 0.270 | 1.1 | 4.4 | 30.7 | 16.1 | <0.001 | 11.9 | <0.001 |
| Apigenin | 132.9 | 182.5 | 524.0 | 140.8 | <0.001 | 19.2 | 55.2 | 9.0 | 11.6 | 0.425 | 0 | 0 | 0 | 0 | 65.1 | <0.001 | |
| Apigenin-7-O-glucoside | 241.9 | 215.8 | 3501.0 | 831.4 | <0.001 | 23.0 | 91.9 | 214.3 | 103.1 | <0.001 | 0 | 0 | 0 | 0 | 45.4 | <0.001 | |
| Santin | 502.4 | 231.3 | 222.8 | 57.31 | <0.001 | 308.7 | 234.4 | 246.0 | 25.4 | 0.241 | 232.4 | 15.2 | 187.9 | 64.5 | 0.011 | 7.3 | 0.001 |
| Total (sum of compounds) | 15,895.4 | 4349.1 | 32,629.0 | 7223.8 | <0.001 | 14,534.0 | 7047.0 | 48,939.8 | 14,818.8 | <0.001 | 4396.5 | 1359.1 | 10,883.2 | 3080.0 | <0.001 | 32.6 | <0.001 |
| Plant Organ | Turkey | p1 | Lithuania | p1 | Acids | Flavonoids | ||
|---|---|---|---|---|---|---|---|---|
| Acids | Flavonoids | Acids | Flavonoids | p2 | p2 | |||
| Flowers | 48.4 a | 51.6 a | 0.501 | 73.8 a | 26.2 a | <0.001 | <0.001 | <0.001 |
| Leaves | 79.9 b | 20.1 b | 0.001 | 88.2 b | 11.8 b | <0.001 | <0.001 | <0.001 |
| Stems | 69.0 b | 31.0 b | <0.001 | 84.4 b | 15.6 b | <0.001 | <0.001 | <0.001 |
| Plant Organ | Turkey | p1 2 | Lithuania | p1 | Acids | Flavonoids | ||
|---|---|---|---|---|---|---|---|---|
| Acids | Flavonoids | Acids | Flavonoids | p2 | p2 | |||
| Flowers | 7456.9 a 1 | 8438.5 a | 0.371 | 23,728.4 a | 8900.6 a | <0.001 | <0.001 | 0.647 |
| Leaves | 11,196.7 b | 3337.3 b | <0.001 | 43,070.4 b | 5869.4 b | <0.001 | <0.001 | 0.002 |
| Stems | 3099.3 c | 1297.1 c | <0.001 | 9185.6 c | 1697.6 c | <0.001 | <0.001 | 0.056 |
| No. | Variables | Inflorescences | Leaves | Stems | |||
|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | ||
| 1 | Neochlorogenic acid | −0.93 | −0.12 | −0.79 | 0.45 | - | - |
| 2 | Chlorogenic acid | −0.96 | −0.12 | −0.92 | 0.21 | −0.93 | −0.08 |
| 3 | 4-O-caffeoylquinic acid | - | - | 0.60 | 0.35 | - | - |
| 4 | 3,4-O-dicaffeoylquinic acid | −0.64 | −0.39 | −0.89 | 0.02 | −0.89 | −0.10 |
| 5 | 3,5-O-dicaffeoylquinic acid | −0.95 | −0.13 | −0.92 | 0.10 | −0.88 | −0.09 |
| 6 | 1,5-O-dicaffeoylquinic acid | −0.91 | 0.03 | −0.85 | −0.10 | −0.86 | −0.24 |
| 7 | 4,5-O-dicaffeoylquinic acid | −0.91 | −0.02 | −0.89 | −0.04 | −0.72 | 0.46 |
| 8 | Caffeic acid | −0.67 | 0.16 | −0.48 | 0.65 | −0.59 | 0.45 |
| 9 | Quercitrin | −0.55 | −0.10 | −0.86 | −0.05 | −0.78 | −0.28 |
| 11 | Quercetin | - | - | 0.55 | −0.34 | 0.52 | −0.40 |
| 12 | Isoquercitrin | 0.58 | −0.26 | - | - | −0.62 | −0.36 |
| 13 | Luteolin | 0.43 | 0.21 | - | - | - | - |
| 14 | Luteolin-7-O-glucoside | −0.59 | −0.59 | −0.75 | −0.44 | - | - |
| 15 | Luteolin-7-O-rutinoside | - | - | −0.87 | −0.07 | - | - |
| 16 | Luteolin-3,7 O-diglucoside | 0.65 | −0.53 | - | - | - | - |
| 17 | Apigenin | −0.78 | 0.30 | - | - | - | - |
| 18 | Apigenin-7-O-glucoside | −0.95 | 0.00 | −0.79 | −0.49 | −0.80 | −0.20 |
| 19 | Santin | 0.64 | −0.36 | - | - | 0.33 | −0.77 |
| Variables | Inflorescences | Leaves | Stems | |||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | |
| Phenolic acids | −0.92 | 0.16 | −0.90 | 0.21 | −0.93 | 0.19 |
| Flavonoids | −0.36 | −0.88 | −0.85 | 0.02 | −0.75 | −0.28 |
| No. | Province, Geographical Region | Latitude (°N) | Longitude (°E) | Elevation (m.a.s.l.) | Habitat |
|---|---|---|---|---|---|
| 1 | Nevşehir, Central Anatolia | 38.6167 | 34.6162 | 954 | Open coniferous woodland |
| 2 | Nevşehir, Central Anatolia | 38.6500 | 34.6495 | 961 | Open coniferous woodland |
| 3 | Nevşehir, Central Anatolia | 38.6833 | 34.7162 | 1012 | Open coniferous woodland |
| 4 | Nevşehir, Central Anatolia | 38.7001 | 34.8663 | 1116 | Open coniferous woodland |
| 5 | Nevşehir; Central Anatolia | 38.7333 | 34.8829 | 1229 | Open coniferous woodland |
| 6 | Nevşehir, Central Anatolia | 38.7833 | 34.7329 | 1287 | Open coniferous woodland |
| 7 | Nevşehir, Central Anatolia | 38.7833 | 34.6162 | 1203 | Open conifer woodland |
| 8 | Gaziantep, Southeast Anatolia | 37.2506 | 37.1492 | 675 | Roadside grassland |
| 9 | Gaziantep, Southeast Anatolia | 37.2501 | 37.1828 | 778 | Roadside grassland |
| 10 | Gaziantep, Southeast Anatolia | 37.2334 | 37.1996 | 1011 | Roadside grassland |
| 11 | Gaziantep, Southeast Anatolia | 37.2333 | 37.2496 | 972 | Calcareous mountain slope |
| 12 | Gaziantep, Southeast Anatolia | 37.2334 | 37.3163 | 933 | Calcareous mountain slope |
| 13 | Gaziantep, Southeast Anatolia | 37.2500 | 37.3163 | 639 | High altitude grassland |
| 14 | Gaziantep, Southeast Anatolia | 37.2667 | 37.3663 | 937 | High altitude grassland |
| 15 | Gaziantep, Southeast Anatolia | 37.2667 | 37.4163 | 946 | High altitude grassland |
| 16 | Gaziantep, Southeast Anatolia | 37.2667 | 37.4663 | 861 | High altitude grassland |
| 17 | Veisiejai, Lazdijai distr. | 54.09871 | 23.72095 | 125 | Dry grassland |
| 18 | Meteliai, Lazdijai distr. | 54.28465 | 23.74717 | 114 | Mixed forest, roadside |
| 19 | Pirčiupiai, Šalčininkai distr. | 54.39087 | 24.95129 | 131 | Pine forest, roadside |
| 20 | Einororys, Alytus distr. | 54.44614 | 24.38971 | 120 | Mesophytic grassland |
| 21 | Geruliai, Alytus distr. | 54.53120 | 24.27056 | 130 | Mesophytic grassland |
| 22 | Gojus, Prienai distr. | 54.56845 | 24.28128 | 96 | Dry grassland |
| 23 | Žuklijai, Trakai distr. | 54.50986 | 24.68252 | 162 | Mesophytic grassland |
| 24 | Pamiškė, Trakai distr. | 54.62607 | 24.51166 | 180 | Mesophytic grassland |
| 25 | Rokantai, Vilnius distr. | 54.73488 | 25.54795 | 180 | Mesophytic grassland |
| 26 | Mickūnai, Vilnius distr. | 54.75274 | 25.55872 | 156 | Pine forest, roadside |
| 27 | Užugriovis, Vilnius distr. | 54.82782 | 25.24644 | 161 | Dry grassland |
| 28 | Bernatonys, Vilnius distr. | 54.90934 | 25.32271 | 160 | Mesophytic grassland |
| 29 | Skardupiai, Šakiai distr. | 54.85567 | 23.03398 | 47 | Mesophytic grassland |
| 30 | Dubingiai, Molėtai distr. | 55.05911 | 25.43509 | 175 | Pine forest, roadside |
| 31 | Apkartai, Molėtai distr. | 55.21658 | 25.64254 | 160 | Dry grassland |
| 32 | Vorėnai, Molėtai distr. | 55.35779 | 25.61012 | 158 | Mesophytic grassland |
| 33 | Mikieriai, Anykščiai dist. | 55.66058 | 25.19964 | 130 | Mesophytic grassland |
| 34 | Svėdasai, Anykščiai distr. | 55.67750 | 25.36114 | 111 | Dry grassland |
| 35 | Maželiai, Rokiškis distr. | 56.03197 | 25.32496 | 99 | Mesophytic grassland |
| 36 | Juodžionys, Biržai distr. | 56.24353 | 24.87915 | 60 | Mesophytic grassland |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radušienė, J.; Karpavičienė, B.; Raudone, L.; Vilkickyte, G.; Çırak, C.; Seyis, F.; Yayla, F.; Marksa, M.; Rimkienė, L.; Ivanauskas, L. Trends in Phenolic Profiles of Achillea millefolium from Different Geographical Gradients. Plants 2023, 12, 746. https://doi.org/10.3390/plants12040746
Radušienė J, Karpavičienė B, Raudone L, Vilkickyte G, Çırak C, Seyis F, Yayla F, Marksa M, Rimkienė L, Ivanauskas L. Trends in Phenolic Profiles of Achillea millefolium from Different Geographical Gradients. Plants. 2023; 12(4):746. https://doi.org/10.3390/plants12040746
Chicago/Turabian StyleRadušienė, Jolita, Birutė Karpavičienė, Lina Raudone, Gabriele Vilkickyte, Cüneyt Çırak, Fatih Seyis, Fatih Yayla, Mindaugas Marksa, Laura Rimkienė, and Liudas Ivanauskas. 2023. "Trends in Phenolic Profiles of Achillea millefolium from Different Geographical Gradients" Plants 12, no. 4: 746. https://doi.org/10.3390/plants12040746
APA StyleRadušienė, J., Karpavičienė, B., Raudone, L., Vilkickyte, G., Çırak, C., Seyis, F., Yayla, F., Marksa, M., Rimkienė, L., & Ivanauskas, L. (2023). Trends in Phenolic Profiles of Achillea millefolium from Different Geographical Gradients. Plants, 12(4), 746. https://doi.org/10.3390/plants12040746