



Agro-Physiological Traits of Kaffir Lime in Response to Pruning and Nitrogen Fertilizer under Mild Shading




Abstract
:1. Introduction
2. Results
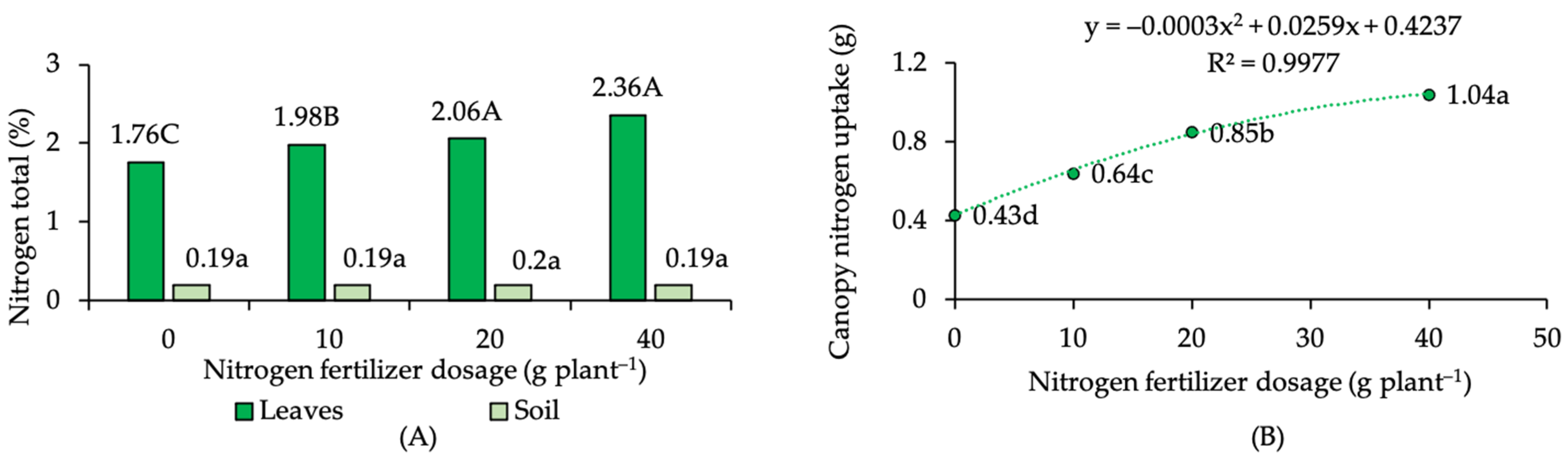
2.1. Nitrogen Status in Soil, Leaves, and Canopy N Uptake
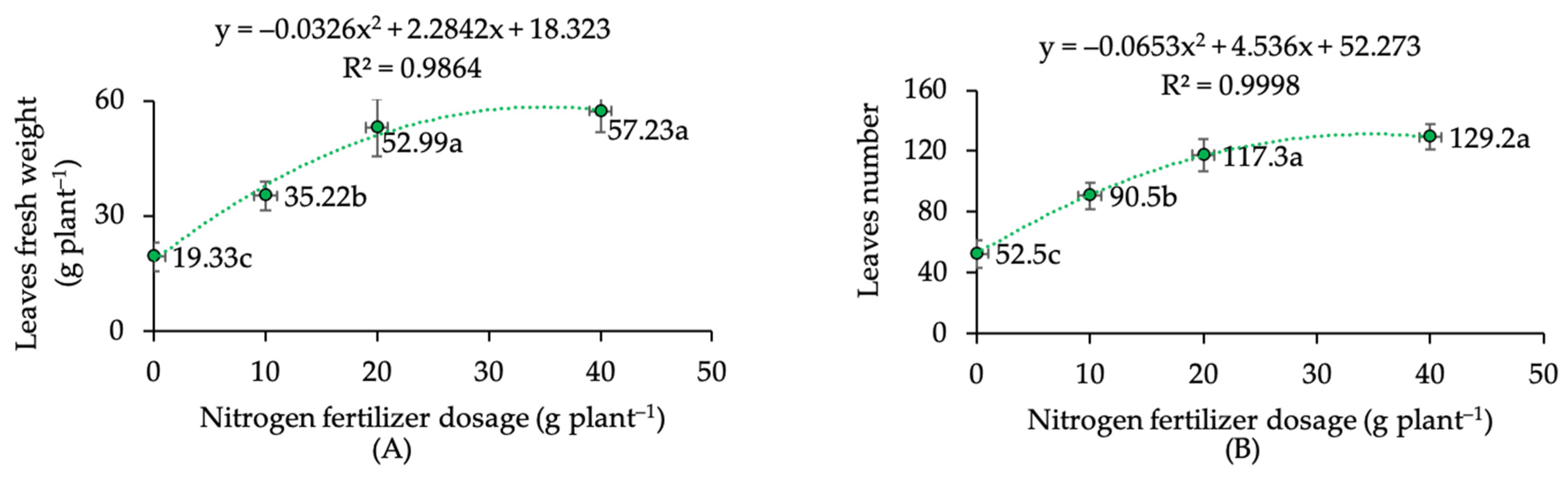
2.2. Growth Performance under Different Pruning Levels and N Dosages
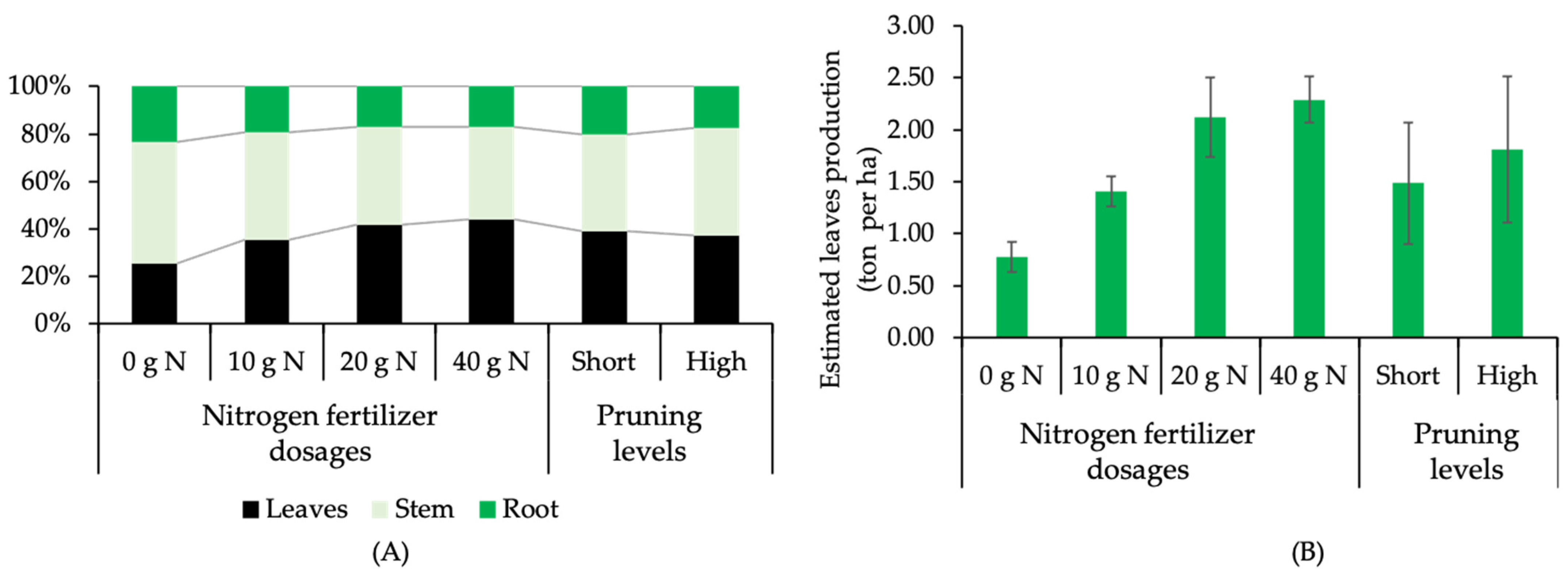
2.3. Plant Production under Different Pruning Levels and N Dosages
2.4. Physiological Response of Kaffir Lime under Different Pruning Levels and N Dosages
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Planting Materials
4.3. Research Procedure
4.4. Measured Variables
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Citrus Fruit Statistics 2015; Market and Policy Analyses of Raw Materials, Horticulture and Tropical (RAMHOT) Products Team Trade and Markets Division; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Nicolosi, E. Origin and Taxonomy. In Citrus Genetics, Breeding and Biotechnology; Cabi: Wallingford, UK, 2007; pp. 19–43. [Google Scholar]
- Qi, L.; Qi, C.J. status quo and development trend of world’s citrus industry. Agric. Outlook 2016, 12, 46–52. [Google Scholar]
- Wu, G.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curk, F.; Ollitrault, F.; Garcia-Lor, A.; Luro, F.; Navarro, L.; Ollitrault, P. Phylogenetic origin of limes and lemons revealed by cytoplasmic and nuclear markers. Ann. Bot. 2016, 11, 565–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabberley, D.J. Citrus (Rutaceae): A review of recent advances in etymology, systematics and medical applications. Blumea Biodivers. Evol. Biogeogr. Plants 2004, 49, 481–498. [Google Scholar] [CrossRef]
- Wongpornchai, S. Kaffir lime leaf. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2012. [Google Scholar]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Sensory evaluation of the quality of kaffir lime (Citrus hystrix DC.) leaves exposed to different postharvest treatments. J. Trop. Crop. Sci. 2021, 8, 71–79. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Production, post- harvest and marketing of kaffir lime (Citrus hystrix DC) in Tulungagung, Indonesia. J. Trop. Crop. Sci. 2019, 6, 138–143. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Morphological evaluation and determination keys of 21 citrus genotypes at seedling stage. Biodiversitas 2021, 22, 1570–1579. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Short communication: Allometric model to estimate bifoliate leaf area and weight of kaffir lime (Citrus hystrix). Biodiversitas 2021, 22, 2815–2820. [Google Scholar] [CrossRef]
- Budiarto, R.; Sholikin, M.M. Kaffir lime essential oil variation in the last fifty years: A meta-analysis of plant origins, plant parts and extraction methods. Horticulturae 2022, 8, 1132. [Google Scholar] [CrossRef]
- Othman, S.N.A.M.; Hassan, M.A.; Nahar, L.; Basar, N.; Jamil, S.; Sarker, S.D. Essential oils from the Malaysian citrus (Rutaceae) medicinal plants. Medicines 2016, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Kooltheat, N.; Kamuthachad, L.; Anthapanya, M.; Samakchan, N.; Sranujit, R.P.; Potup, P.; Ferrante, A.; Usuwanthim, K. Kaffir lime leaves extract inhibits biofilm formation by Streptococcus mutans. Nutrition 2016, 32, 486–490. [Google Scholar] [CrossRef]
- Ansori, A.N.M.; Supriyadi, A.P.; Kartjito, M.V.; Rizqi, F.; Adrianto, H. Biolarvacidal effectivities of polar and non-polar extract fraction from kaffir lime (Citrus hystrix) leaf against 3rd instar larvae of Aedes aegypti. J. Biol. Eng. Res. Rev. 2015, 2, 14–17. [Google Scholar]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Agronomical and physiological characters of kaffir lime (Citrus hystrix DC) seedling under artificial shading and pruning. Emir. J. Food Agric. 2019, 31, 222–230. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Shoot manipulations improve flushing and flowering of mandarin citrus in Indonesia. J. Appl. Hortic. 2018, 20, 112–118. [Google Scholar] [CrossRef]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.A.; Laghari, A.H.; Bhabhan, G.M.; Talpur, K.H.; Bhutto, T.A.; Wahocho, S.A.; Lashari, A.A. Role of nitrogen for plant growth and development: A review. Adv. Environ. Biol. 2016, 10, 209–218. [Google Scholar]
- Muñoz-Huerta, R.F.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R.V. A review of methods for sensing the nitrogen status in plants: Advantages, disadvantages and recent advances. Sensors 2013, 13, 10823–10843. [Google Scholar] [CrossRef]
- Liao, L.; Dong, T.; Qiu, X.; Rong, Y.; Wang, Z.; Zhu, J. Nitrogen nutrition is a key modulator of the sugar and organic acid content in citrus fruit. PLoS ONE 2019, 14, e0223356. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen nutrition of fruit trees to reconcile productivity and environmental concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Boughalleb, F.; Mahmoud, M.; Hajlaoui, H. Response of young citrus trees to NPK fertilization under greenhouse and field conditions. Agric. J. 2011, 6, 66–73. [Google Scholar] [CrossRef]
- Efendi, D.; Budiarto, R.; Poerwanto, R.; Santosa, E.; Agusta, A. Relationship among agroclimatic variables, soil and leaves nutrient status with the yield and main composition of kaffir lime (Citrus hystrix DC) leaves essential oil. Metabolites 2021, 11, 260. [Google Scholar] [CrossRef] [PubMed]
- Fertilizer Recommendations for Citrus Plants (Rekomendasi Pemupukan untuk Tanaman Jeruk). Available online: http://balitjestro.litbang.pertanian.go.id/rekomendasi-pemupukan-untuk-tanaman-jeruk/ (accessed on 18 December 2021).
- Morales, P.; Davies, F.S. Pruning and skirting affect canopy microclimate, yields and fruit quality of Orlando Tangelo. Hortscience 2000, 35, 30–35. [Google Scholar] [CrossRef]
- Joubert, F.J.; Du Plessis, M.H.; Stassen, P.J.C. Pruning strategies to alleviate overcrowding in higher density citrus orchards. J. Appl. Hortic. 2000, 2, 1–5. [Google Scholar] [CrossRef]
- Gilman, E.F.; Black, R.J. Pruning Landscape Trees and Shrubs; University of Florida: Gainesville, FL, USA, 2011. [Google Scholar]
- Pruning Citrus. Available online: http://www.ucanr.edu/sites/placernevadasmallfarms/files/134946.pdf (accessed on 22 December 2022).
- Eissenstat, D.M.; Duncan, L.W. Root growth and carbohydrate responses in bearing citrus trees following partial canopy removal. Tree Physiol. 1992, 10, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.L. Tree size control hedging and topping citrus in high-density plantings. Proc. Fla. State Hort. Soc. 1978, 91, 43–46. [Google Scholar]
- Goldschmidt, E.E. Carbohydrate supply as a critical factor for citrus fruit development and productivity. Hortscience 1999, 34, 1020–1024. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.D.; Singh, M.; Rawls, E.K.; Taylor, J.B., Jr. Defoliation of citrus trees by diquat. Proc. Fla. State Hort. Soc. 2007, 120, 120–126. [Google Scholar]
- Aliyah, M.; Susanto, S.; Sukma, D.; Ardie, S.W. Performance improvement of young pummelo citrus (Citrus maxima (Burm.) Merr.) by strangulation application and pinching. Asian J. Agric. Res. 2015, 9, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Dashora, L.K.; Bhatnagar, P.; Singh, B. Impact of pruning on rejuvenation of declining Nagpur mandarin (Citrus reticulata Blanco.) orchard. Indian J. Agrofor. 2016, 18, 53–57. [Google Scholar]
- Yuan, R.; Alferez, F.; Kostenyuk, I.; Singh, S.; Syvertsen, J.P.; Burns, J.K. Partial defoliation can decrease average leaf size but has little effect on orange tree growth, fruit yield and juice quality. Hortscience 2005, 40, 2011–2015. [Google Scholar] [CrossRef] [Green Version]
- Ingle, I.V.; Athawale, R.B.; Tayde, G.S.; Pakhare, G.B. Effect of severity and time of pruning on flower type fruit set and fruit retention in old acid lime trees (Citrus aurantifolia). Agric. Sci. Digest. 2001, 21, 65–66. [Google Scholar]
- Poerwanto, R.; Susila, A.D. Teknologi Hortikutura; IPB Press: Bogor, Indonesia, 2014. [Google Scholar]
- Quaggio, J.A.; Cantarella, H.; Van Raij, B. Phosphorus and potassium soil test and nitrogen leaf analysis as a base for citrus fertilization. Nutr. Cycl. Agroecosyst. 1998, 52, 67–74. [Google Scholar] [CrossRef]
- Obreza, T.A.; Alva, A.K.; Hanlon, E.A.; Rouse, R.E. Citrus Grove Leaf Tissue and Soil Testing: Sampling, Analysis, and Interpretation; Fact Sheet; Florida Cooperative Extension Service: Gainesville, FL, USA, 1992; Volume 115, pp. 1–5. [Google Scholar]
- Bennett, W.F. Nutrient Deficiencies and Toxicities in Crop Plants; APS Press: St. Paul, MN, USA, 1993; p. 202. [Google Scholar]
- Mills, H.A.; Benton-Jones, J., Jr. Plant Analysis Handbook II; Micromacro Publishing: Athens, GA, USA, 1996; p. 422. [Google Scholar]
- Srivastava, A.K.; Singh, S. Leaf and soil nutrient guide in citrus: A review. Agric. Rev. 2004, 25, 235–251. [Google Scholar]
- Raveh, E. Citrus leaf nutrient status: A critical evaluation of guidelines for optimal yield in Israel. J. Plant Nutr. Soil Sci. 2013, 176, 420–428. [Google Scholar] [CrossRef]
- Bathurst, A.C. New method for estimating the fertilizer requirements of citrus trees. Farming Soc. Afr. 1943, 18, 323–327. [Google Scholar]
- Chapman, H.D. Citrus leaf analysis. Nutrient deficiencies, excesses and fertilizer requirements of soil indicated by diagnostic aid. Clif. Agric. 1949, 3, 10–14. [Google Scholar]
- Reuther, W.; Smith, P.F. Tissue analysis as an aid in evaluating the nutritional status of citrus trees. Proc. Rio Gd. Val. Hort. Inst. 1951, 5, 34–35. [Google Scholar]
- Embleton, T.W.; Jones, W.W.; Labanauskas, C.; Reuther, W. Leaf analysis as a diagnostic tool and guide to fertilization. In The Citrus Industry; Reuther, W., Ed.; University of California Press: Berkeley, CA, USA, 1973; Volume III, pp. 183–210. [Google Scholar]
- Alva, A.K.; Paramasivam, S.; Obreza, T.A.; Schumann, A.W. Nitrogen best management practice for citrus trees: I. Fruit yield, quality, and leaf nutritional status. Sci. Hort. 2006, 107, 233–244. [Google Scholar] [CrossRef]
- Obreza, T.A.; Zekri, M.; Hanlon, E.A. Soil and leaf tissue testing. In Nutrition of Florida Citrus Trees, 2nd ed.; Obreza, T.A., Morgan, K.T., Eds.; Soil and Water Science Department, University of Florida Press: Gainesville, FL, USA, 2011; pp. 24–32. [Google Scholar]
- Bondada, B.R.; Syvertsen, J.P.; Albrigo, L.G. urea nitrogen uptake by citrus leaves. Hortscience 2001, 36, 1061–1065. [Google Scholar] [CrossRef] [Green Version]
- Kohli, R.R.; Srivastava, A.K.; Huchche, A.D.; Dass, H.C.; Ram, L.; Singh, S. Diagnosis of leaf nutrient level for optimum productivity of Citrus reticulata Blanco grown in black clay soils under a sub-humid tropical climate. Trop. Agric. Res. Ext. 1998, 1, 81–86. [Google Scholar]
- Fahl, J.I.; Carelli, M.L.C.; Vega, J.; Magalhães, A.C. Nitrogen and irradiance levels affecting net photosynthesis and growth of young coffee plants (Coffea arabica L.). J. Hortic. Sci. 1994, 69, 161–169. [Google Scholar] [CrossRef]
- Lin, M.T.; Chen, A.M.; Lin, T.S.; Kuan, C.S.; Lee, C.L.; Yang, W.J. Prevention of natural flowering in pineapple (Ananas comosus) by shading and urea application. Hort Environ. Biotechnol. 2015, 56, 9–16. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Islam, M.S.; Hossain, M.M.; Hossain, T.; Miah, M.G. Effects of shade and nitrogen levels on quality Bangladhonia (leafy green culinary of corriander relatives) production. Bangladesh J. Agric. Res. 2009, 34, 205–213. [Google Scholar] [CrossRef]
- McCauley, A.; Jacobsen, J. Plant Nutrient Functions and Deficiency and Toxicity Symptoms; Montana State University Extension: Bozeman, MT, USA, 2011. [Google Scholar]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Preliminary study on antioxidant and antibacterial activity of kaffir lime (Citrus hystrix DC) leaf essential oil. Appl. Res. Sci. Technol. 2021, 1, 58–65. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. The effects of preharvest mild shading on the quality and production of essential oil from kaffir lime leaves (Citrus hystrix). J. Trop. Crop. Sci. 2022, 9, 15–21. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Comparative analysis and correlation of young and mature kaffir lime (Citrus hystrix DC) leaf characteristics. Int. J. Plant Biol. 2022, 13, 23. [Google Scholar] [CrossRef]
- Barker, V.A.; Pilbeam, D.J. Introduction. In Handbook of Plant Nutrition; Barker, A.V., Pilbeal, D.J., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Huang, W.T.; Xie, Y.Z.; Chen, X.F.; Zhang, J.; Chen, H.H.; Ye, X.; Guo, J.; Yang, L.T.; Chen, L.S. Growth, mineral nutrients, photosynthesis and related physiological parameters of Citrus in response to nitrogen deficiency. Agronomy 2021, 11, 1859. [Google Scholar] [CrossRef]
- Bondada, B.R.; Syvertsen, J.P. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure of citrus leaves under different nitrogen status. Tree Physiol. 2003, 23, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, D.A.; Gamon, J.A. Relationship between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NTS | NLC | CNU | RGR | PH | SN | LN | LFW | PFW | PR | SC | TR | CA | CB | CT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NLC | 0.05 | ||||||||||||||
| CNU | 0.28 | 0.97 | |||||||||||||
| RGR | 0.42 | 0.93 | 0.98 | ||||||||||||
| PH | 0.57 | 0.84 | 0.94 | 0.97 | |||||||||||
| SN | 0.15 | 0.98 | 0.98 | 0.96 | 0.87 | ||||||||||
| LN | 0.39 | 0.93 | 0.98 | 0.99 | 0.94 | 0.97 | |||||||||
| LFW | 0.61 | 0.82 | 0.92 | 0.97 | 0.97 | 0.87 | 0.97 | ||||||||
| PFW | 0.44 | 0.92 | 0.98 | 0.99 | 0.97 | 0.95 | 0.99 | 0.98 | |||||||
| PR | 0.54 | 0.82 | 0.92 | 0.94 | 0.99 | 0.84 | 0.90 | 0.93 | 0.94 | ||||||
| SC | 0.40 | 0.27 | 0.33 | 0.43 | 0.30 | 0.40 | 0.50 | 0.53 | 0.44 | 0.17 | |||||
| TR | 0.53 | −0.79 | −0.65 | −0.51 | −0.40 | −0.71 | −0.52 | −0.30 | −0.50 | −0.43 | 0.19 | ||||
| CA | 0.07 | 0.98 | 0.97 | 0.91 | 0.86 | 0.96 | 0.90 | 0.80 | 0.91 | 0.86 | 0.13 | −0.81 | |||
| CB | 0.05 | 0.98 | 0.96 | 0.91 | 0.85 | 0.96 | 0.89 | 0.79 | 0.90 | 0.85 | 0.13 | −0.82 | 0.99 | ||
| CT | 0.06 | 0.98 | 0.97 | 0.91 | 0.86 | 0.96 | 0.90 | 0.80 | 0.90 | 0.86 | 0.13 | −0.81 | 1.00 | 0.99 | |
| ANT | −0.77 | 0.58 | 0.37 | 0.24 | 0.04 | 0.50 | 0.28 | 0.02 | 0.22 | 0.04 | −0.04 | −0.89 | 0.53 | 0.55 | 0.54 |
| Treatment | RGR (g week−1) | Plant Height (cm) | Shoot Number | Leaf Numbers |
|---|---|---|---|---|
| Nitrogen fertilizer (N) factor | ||||
| 0 g N | 1.71 ± 0.27 c | 53.75 ± 14.75 b | 6.50 ± 1.05 b | 52.50 ± 9.07 c |
| 10 g N | 2.39 ± 0.22 b | 58.75 ± 13.65 b | 7.50 ± 1.38a b | 90.50 ± 8.55 b |
| 20 g N | 3.15 ± 0.43 a | 79.13 ± 24.91 a | 7.83 ± 1.72 a | 117.33 ± 10.31 a |
| 40 g N | 3.40 ± 0.38 a | 79.29 ± 23.27 a | 8.67 ± 2.34 a | 129.17 ± 8.40 a |
| Pruning (P) factor | ||||
| SP (10 cm) | 2.42 ± 0.65 b | 51.87 ± 8.95 b | 8.92 ± 1.56 a | 92.92 ± 32.54 a |
| HP (30 cm) | 2.90 ± 0.78 a | 83.59 ± 19.41 a | 6.33 ± 0.65 b | 101.83 ± 30.69 a |
| N*P | Ns | Ns | Ns | Ns |
| Treatment | Fresh Weight (g) | |||
|---|---|---|---|---|
| Plant | Leaves | Stem | Root | |
| Nitrogen-fertilizer (N) factor | ||||
| 0 g N | 76.36 ± 13.18 c | 19.33 ± 3.65 c | 39.29 ± 8. 21 c | 17. 75 ± 1.73 c |
| 10 g N | 99.10 ± 11.74 b | 35.22 ± 3.67 b | 44.69 ± 7. 53 b | 19. 18 ± 1. 72 bc |
| 20 g N | 124.51 ± 20.15 a | 52. 99 ± 9. 46 a | 51. 94 ± 9. 82 a | 21.31 ± 1. 83 ab |
| 40 g N | 131.53 ± 16.56 a | 57.23 ± 5. 59 a | 50. 21 ± 10.91 a | 22. 37 ± 1.43 a |
| Pruning (P) factor | ||||
| SP (10 cm) | 94.71 ± 20.98 b | 37. 12 ± 14. 67 b | 38. 46 ± 4. 44 b | 19. 13 ± 2. 36 b |
| HP (30 cm) | 121.03 ± 25.94 a | 45. 26 ± 17. 66 a | 54.61 ± 6.80 a | 21. 16 ± 2. 10 a |
| N*P | Ns | Ns | Ns | Ns |
| Treatment | Pn | Tr | Sc | WUE | LUE |
|---|---|---|---|---|---|
| Nitrogen-fertilizer (N) factor | |||||
| 0 g N | 17.52 ± 1.19 b | 6.54 ± 1.56 | 0.371 ± 0.09 | 2.68 ± 0.76 | 0.58 ± 1.56 |
| 10 g N | 17.63 ± 1.22 b | 6.50 ± 1.85 | 0.383 ± 0.12 | 2.74 ± 0.66 | 0.74 ± 1.85 |
| 20 g N | 20.03 ± 1.13 a | 6.59 ± 1.39 | 0.381 ± 0.09 | 3.04 ± 0.81 | 0.92 ± 1.39 |
| 40 g N | 20.20 ± 0.99 a | 6.25 ± 0.96 | 0.376 ± 0.06 | 3.23 ± 1.03 | 0.98 ± 0.96 |
| Pruning (P) factor | |||||
| H10 | 18.69 ± 1.81 | 6.70 ± 1.64 | 0.397 ± 0.10 a | 2.81 ± 1. 10 | 0.71 ± 1.64 |
| H30 | 18.99 ± 1.59 | 6.24 ± 1.22 | 0.358 ± 0.08 b | 3.04 ± 1. 30 | 0.90 ± 1.22 |
| N*P | Ns | Ns | Ns | Ns | Ns |
| Treatment | Chlorophyll α (mg g−1) | Chlorophyll β (mg g−1) | Chlorophyll Total (mg g−1) | Anthocyanin (mg 100 g−1) |
|---|---|---|---|---|
| 0 g N | 0.319 ± 0.12 d | 0.122 ± 0.06 d | 0.441 ± 0.17 d | 0.063 ± 0.04 a |
| 10 g N | 0.618 ± 0.08 c | 0.229 ± 0.02 c | 0.846 ± 0.10 c | 0.077 ± 0.01 a |
| 20 g N | 0.953 ± 0.06 b | 0.337 ± 0.02 b | 1292 ± 0.09 b | 0.051 ± 0.01 a |
| 40 g N | 1.687 ± 0.12 a | 0.603 ± 0.04 a | 2.291 ± 0.16 a | 0.088 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Agro-Physiological Traits of Kaffir Lime in Response to Pruning and Nitrogen Fertilizer under Mild Shading. Plants 2023, 12, 1155. https://doi.org/10.3390/plants12051155
Budiarto R, Poerwanto R, Santosa E, Efendi D, Agusta A. Agro-Physiological Traits of Kaffir Lime in Response to Pruning and Nitrogen Fertilizer under Mild Shading. Plants. 2023; 12(5):1155. https://doi.org/10.3390/plants12051155
Chicago/Turabian StyleBudiarto, Rahmat, Roedhy Poerwanto, Edi Santosa, Darda Efendi, and Andria Agusta. 2023. "Agro-Physiological Traits of Kaffir Lime in Response to Pruning and Nitrogen Fertilizer under Mild Shading" Plants 12, no. 5: 1155. https://doi.org/10.3390/plants12051155
APA StyleBudiarto, R., Poerwanto, R., Santosa, E., Efendi, D., & Agusta, A. (2023). Agro-Physiological Traits of Kaffir Lime in Response to Pruning and Nitrogen Fertilizer under Mild Shading. Plants, 12(5), 1155. https://doi.org/10.3390/plants12051155