
Different Responses of Growing Season Ecosystem CO2 Fluxes to Rain Addition in a Desert Ecosystem
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
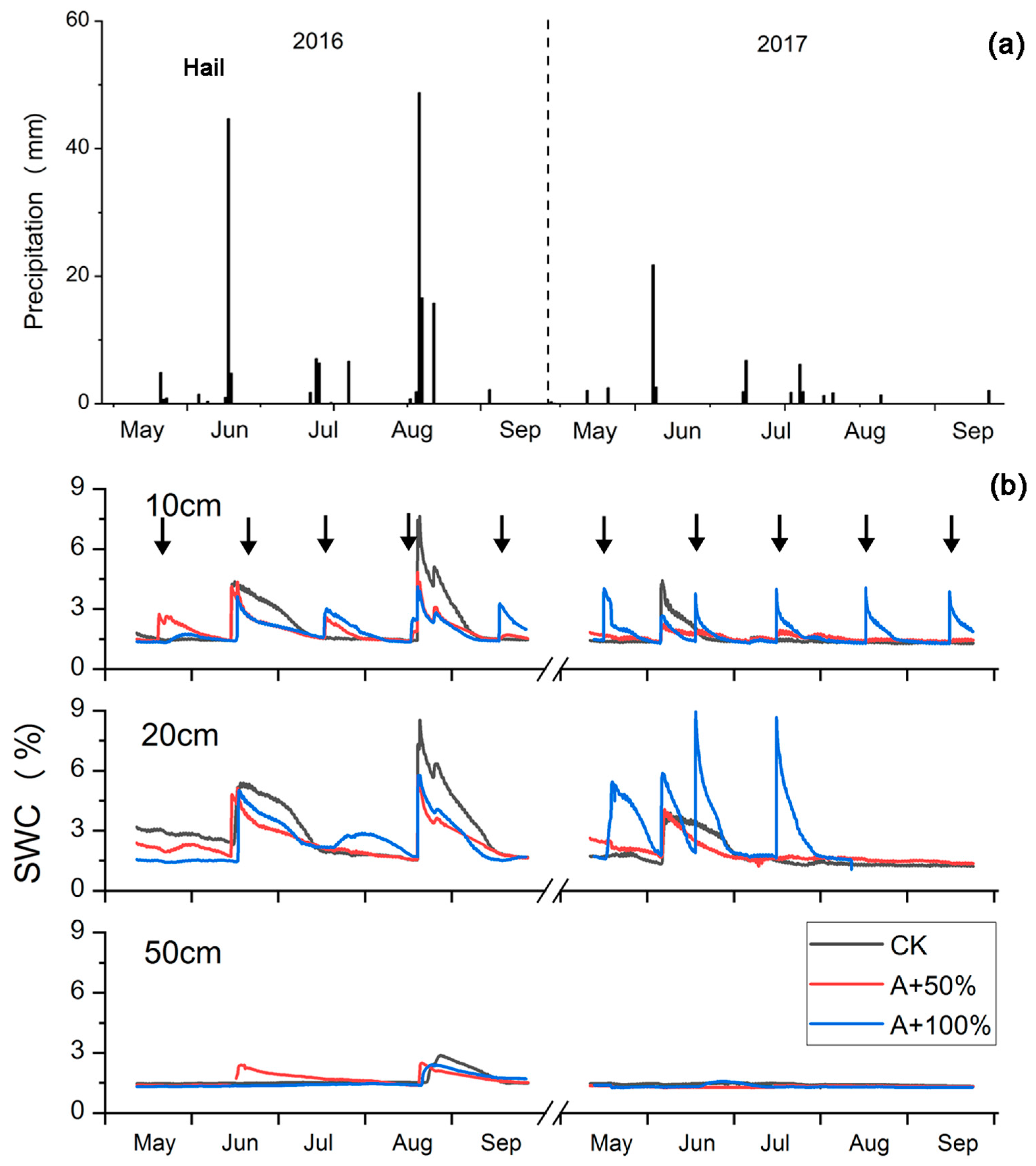
2.1. Rain Addition Treatments and SWC Changes
2.2. Response of Ecosystem CO2 Fluxes to Rain Addition
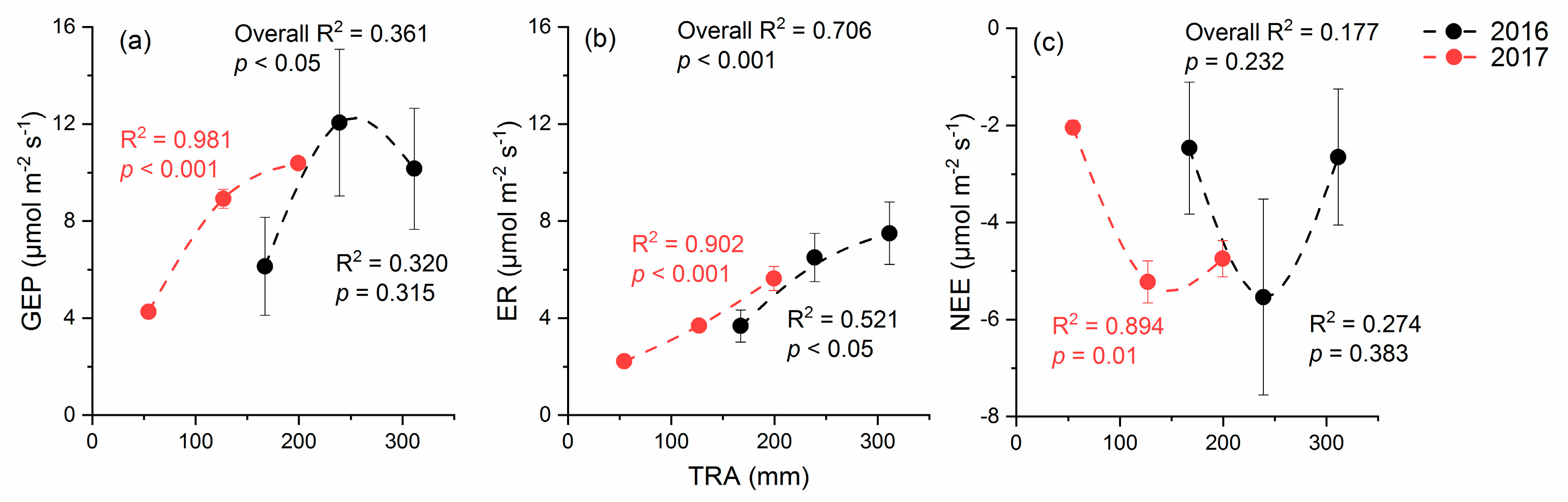
2.3. Response of CO2 Fluxes to Total Rainfall Amount (TRA)
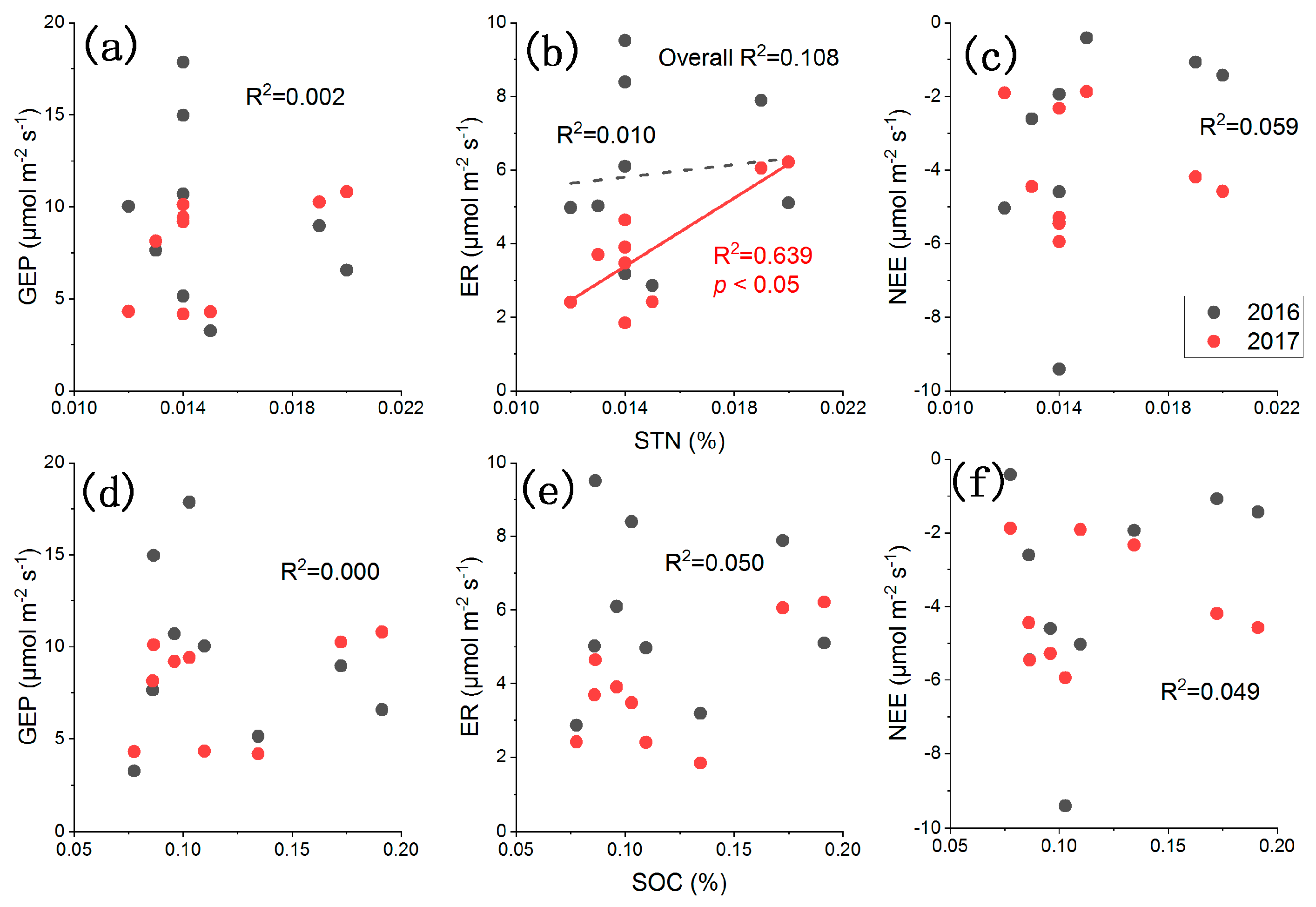
2.4. Influences of Environmental Factors on CO2 Fluxes
3. Discussion
3.1. Differential Response Patterns of CO2 Fluxes
3.2. Rain Addition Enhanced NEE of the Desert Ecosystem
3.3. Implications for the Carbon Cycling of Desert Ecosystems
4. Materials and Methods
4.1. Study Area
4.2. Experimental Design
4.3. Ecosystem CO2 Flux Measurements
4.4. Soil Property Measurements
4.5. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R. Carbon sequestration in dryland ecosystems. Environ. Manag. 2004, 33, 528–544. [Google Scholar] [CrossRef]
- Stone, R. Ecosystems—Have desert researchers discovered a hidden loop in the carbon cycle? Science 2008, 320, 1409–1410. [Google Scholar] [CrossRef] [PubMed]
- Ahlstrom, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasoni, R.L.; Smith, S.D.; Arnone, J.A. Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2. Glob. Chang. Biol. 2005, 11, 749–756. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Fenstermaker, L.F.; Iii, J.A.A. Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem. Glob. Chang. Biol. 2008, 14, 1475–1487. [Google Scholar] [CrossRef]
- Xie, J.; Li, Y.; Zhai, C.; Li, C.; Lan, Z. CO2 absorption by alkaline soils and its implication to the global carbon cycle. Environ. Geol. 2009, 56, 953–961. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Belnap, J.; Marion, G. On carbon sequestration in desert ecosystems. Glob. Chang. Biol. 2009, 15, 1488–1490. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Zhang, X.; Zwiers, F.W.; Hegerl, G.C.; Lambert, F.H.; Gillett, N.P.; Solomon, S.; Stott, P.A.; Nozawa, T. Detection of human influence on twentieth-century precipitation trends. Nature 2007, 448, 461–465. [Google Scholar] [CrossRef]
- Li, Y.; Huang, J.; Ji, M.; Ran, J. Dryland Expansion in Northern China from 1948 to 2008. Adv. Atmos. Sci. 2015, 32, 870–876. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.; Liu, L.; Jia, R.; Yang, H.; Li, G.; Wei, Y. Seasonal variation of carbon exchange from a revegetation area in a Chinese desert. Agric. For. Meteorol. 2012, 156, 134–142. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.; Qin, D.; Wu, J.; Gao, R.; Song, L. Changes in Mean and Extreme Temperature and Precipitation over the Arid Region of Northwestern China: Observation and Projection. Adv. Atmos. Sci. 2017, 34, 9–25. [Google Scholar] [CrossRef]
- Rodríguez-Iturbe, I.; Porporato, A. Ecohydrology of Water-Controlled Ecosystems: Soil Moisture and Plant Dynamics; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Fay, P.A.; Kaufman, D.M.; Nippert, J.B.; Carlisle, J.D.; Harper, C.W. Changes in grassland ecosystem function due to extreme rainfall events: Implications for responses to climate change. Glob. Chang. Biol. 2008, 14, 1600–1608. [Google Scholar] [CrossRef]
- Felton, A.J.; Slette, I.J.; Smith, M.D.; Knapp, A.K. Precipitation amount and event size interact to reduce ecosystem functioning during dry years in a mesic grassland. Glob. Chang. Biol. 2020, 26, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of More Extreme Precipitation Regimes for Terrestrial Ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Loik, M.E.; Schwinning, S.; Williams, D.G.; Fay, P.A.; Haddad, B.M.; Harte, J.; Huxman, T.E.; Knapp, A.K.; Lin, G.; et al. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience 2003, 53, 941–952. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, K.R.; von Fischer, J.C.; Muscha, J.M.; Petersen, M.K.; Knapp, A.K. Contrasting above- and belowground sensitivity of three Great Plains grasslands to altered rainfall regimes. Glob. Chang. Biol. 2015, 21, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Zeppel, M.J.B.; Wilks, J.V.; Lewis, J.D. Impacts of extreme precipitation and seasonal changes in precipitation on plants. Biogeosciences 2014, 11, 3083–3093. [Google Scholar] [CrossRef] [Green Version]
- Beier, C.; Beierkuhnlein, C.; Wohlgemuth, T.; Penuelas, J.; Emmett, B.; Körner, C.; de Boeck, H.; Christensen, J.H.; Leuzinger, S.; Janssens, I.A.; et al. Precipitation manipulation experiments—Challenges and recommendations for the future. Ecol. Lett. 2012, 15, 899–911. [Google Scholar] [CrossRef]
- Chen, S.; Lin, G.; Huang, J.; Jenerette, G.D. Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Glob. Chang. Biol. 2009, 15, 2450–2461. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung M Richardson, A.D.; Papale, D.; Piao, S.; Schulze, E.D.; Wingate, L.; Matteucci, G. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef] [Green Version]
- Ru, J.; Zhou, Y.; Hui, D.; Zheng, M.; Wan, S. Shifts of growing-season precipitation peaks decrease soil respiration in a semiarid grassland. Glob. Chang. Biol. 2018, 24, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L.; Cable, J.; Potts, D.; Ignace, D.; Barron-Gafford, G.; Griffith, A.; Alpert, H.; Van Gestel, N.; Robertson, T.; Huxman, T.E.; et al. Effects of an increase in summer precipitation on leaf, soil, and ecosystem fluxes of CO2 and H2O in a sotol grassland in Big Bend National Park, Texas. Oecologia 2007, 151, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Hamerlynck, E.P.; Scott, R.L.; Stone, J.J. Soil moisture and ecosystem function responses of desert grassland varying in vegetative cover to a saturating precipitation pulse. Ecohydrology 2012, 5, 297–305. [Google Scholar] [CrossRef]
- Snyder, K.A.; Donovan, L.A.; James, J.J.; Tiller, R.L.; Richards, J.H. Extensive summer water pulses do not necessarily lead to canopy growth of Great Basin and northern Mojave Desert shrubs. Oecologia 2004, 141, 325–334. [Google Scholar] [CrossRef]
- Huxman, T.E.; Cable, J.M.; Ignace, D.D.; Eilts, J.A.; English, N.B.; Weltzin, J.; Williams, D.G. Response of net ecosystem gas exchange to a simulated precipitation pulse in a semi-arid grassland: The role of native versus non-native grasses and soil texture. Oecologia 2004, 141, 295–305. [Google Scholar] [CrossRef]
- Potts, D.L.; Huxman, T.E.; Cable, J.M.; English, N.B.; Ignace, D.D.; Eilts, J.A.; Mason, M.J.; Weltzin, J.F.; Williams, D.G. Antecedent moisture and seasonal precipitation influence the response of canopy-scale carbon and water exchange to rainfall pulses in a semi-arid grassland. New Phytol. 2006, 170, 849–860. [Google Scholar] [CrossRef]
- Cable, J.M.; Ogle, K.; Williams, D.G.; Weltzin, J.F.; Huxman, T.E. Soil texture drives responses of soil respiration to precipitation pulses in the Sonoran Desert: Implications for climate change. Ecosystems 2008, 11, 961–979. [Google Scholar] [CrossRef]
- Huang, G.; Li, Y.; Padilla, F.M. Ephemeral plants mediate responses of ecosystem carbon exchange to increased precipitation in a temperate desert. Agric. For. Meteorol. 2015, 201, 141–152. [Google Scholar] [CrossRef]
- Knapp, A.K.; Ciais, P.; Smith, M.D. Reconciling inconsistencies in precipitation–productivity relationships: Implications for climate change. New Phytol. 2017, 214, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Burkett, V.R.; Wilcox, D.A.; Stottlemyer, R.; Barrow, W.; Fagre, D.; Baron, J.; Price, J.; Nielsen, J.L.; Allen, C.D.; Peterson, D.L.; et al. Nonlinear dynamics in ecosystem response to climatic change: Case studies and policy implications. Ecol. Complex. 2005, 2, 357–394. [Google Scholar] [CrossRef] [Green Version]
- Flombaum, P.; Yahdjian, L.; Sala, O.E. Global-change drivers of ecosystem functioning modulated by natural variability and saturating responses. Glob. Chang. Biol. 2017, 23, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kang, X.; Biederman, J.A.; Wang, W.; Qian, R.; Zheng, Z.; Zhang, B.; Ran, Q.; Xu, C.; Liu, W.; et al. Nonlinear carbon cycling responses to precipitation variability in a semiarid grassland. Sci. Total Environ. 2021, 781, 147062. [Google Scholar] [CrossRef]
- Zhang, F.; Quan, Q.; Ma, F.; Tian, D.; Zhou, Q.; Niu, S. Differential responses of ecosystem carbon flux components to experimental precipitation gradient in an alpine meadow. Funct. Ecol. 2019, 33, 889–900. [Google Scholar] [CrossRef]
- Zhou, X.; Weng, E.; Luo, Y. Modeling patterns of nonlinearity in ecosystem responses to temperature, CO2, and precipitation changes. Ecol. Appl. 2008, 18, 453–466. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P.; Fridley, J.D.; Askew, A.P.; Thompson, K.; Hodgson, J.G.; Bennett, C.R. Long-term resistance to simulated climate change in an infertile grassland. Proc. Natl. Acad. Sci. USA 2008, 105, 10028–10032. [Google Scholar] [CrossRef] [Green Version]
- Yahdjian, L.; Sala, O.E. Vegetation structure constrains primary production response to water availability in the Patagonian steppe. Ecology 2006, 87, 952–962. [Google Scholar] [CrossRef]
- Harper, C.W.; Blair, J.M.; Fay, P.A.; Knapp, A.K.; Carlisle, J.D. Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem. Glob. Chang. Biol. 2005, 11, 322–334. [Google Scholar] [CrossRef]
- Miao, Y.; Han, H.; Du, Y.; Zhang, Q.; Jiang, L.; Hui, D.; Wan, S. Nonlinear responses of soil respiration to precipitation changes in a semiarid temperate steppe. Sci. Rep. 2017, 7, 45782. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Chen, S.; Wu, B.; Zhu, Y.; Zhou, Y.; Li, Y.; Cao, Y.; Lu, Q.; Lin, G. Vegetation cover and rain timing co-regulate the responses of soil CO2 efflux to rain increase in an arid desert ecosystem. Soil Biol. Biochem. 2012, 49, 114–123. [Google Scholar] [CrossRef]
- Pan, X.; Shen, G.; Chen, P. A preliminary research on taxonomy and systematics of genus Nitraria. Acta Bot. Yunanica 1999, 21, 287–295. (In Chinese) [Google Scholar]
- Li, Q.; Xu, J.; Li, H.; Wang, S.; Yan, X.; Xin, Z.; Jiang, Z.; Wang, L.; Jia, Z. Effects of aspect on clonal reproduction and biomass allocation of layering modules of Nitraria tangutorum in nebkha dunes. PLoS ONE 2013, 8, e79927. [Google Scholar] [CrossRef] [PubMed]
- Lv, J. Brief on exploitation and utility of Nitraria in Chaidamu. Sci. Technol. Qinghai Agric. For. 2005, 4, 38–40. (In Chinese) [Google Scholar]
- Ren, J.; Tao, L. A numerical taxonomy of the genus Nitraria from Gansu provence, China. Acta Bot. Boreali-Occident. Sin. 2003, 23, 572–576. (In Chinese) [Google Scholar]
- He, J.; Bao, F.; Wu, B.; Zhang, J.; Jia, Z. Photosynthetic acclimation of a desert species Nitraria tangutorum to long term rain addition in north-western China. Int. J. Agric. Biol. 2019, 22, 1050–1058. [Google Scholar]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernandez, R.J. Modifying the “pulse reserve” paradigm for deserts of North America: Precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Zhang, J.; Gu, L.; Bao, F.; Cao, Y.; Hao, Y.; He, J.; Meng, P. Nitrogen control of 13C enrichment in heterotrophic organs relative to leaves in a landscape-building desert plant species. Biogeosciences 2015, 12, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef]
- Ren, H.; Xu, Z.; Isbell, F.; Huang, J.; Han, X.; Wan, S.; Chen, S.; Wang, R.; Zeng, D.; Jiang, Y.; et al. Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation. Ecol. Monogr. 2017, 87, 457–469. [Google Scholar] [CrossRef]
- Hooper, D.U.; Johnson, L. Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation. Biogeochemistry 1999, 46, 247–293. [Google Scholar] [CrossRef]
- Maestre, F.T.; Reynolds, J.F. Amount or pattern? Grassland responses to the heterogeneity and availability of two key resources. Ecology 2007, 88, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jia, Z.; Wu, B.; Lu, Q.; Yao, B. The role of increased precipitation in promoting branch and leaf growth of Nitraria tangutorum. For. Res. 2012, 25, 626–631. (In Chinese) [Google Scholar]
- Gao, Y.; Xu, X.; Ding, J.; Bao, F.; De Costa, Y.G.; Zhuang, W.; Wu, B. The responses to long-term water addition of soil bacterial, archaeal and fungal communities in a desert ecosystem. Microorganisms 2021, 9, 981. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, Q.; Wu, B.; Zhu, Y.; Li, Y. Responses of Nitraria tangutorum branch and leaf growth to simulated rainfall. For. Res. 2012, 25, 130–137. (In Chinese) [Google Scholar]
- Niu, S.; Wu, M.; Han, Y.; Xia, J.; Li, L.; Wan, S. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. New Phytol. 2008, 177, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.W.; Acosta-Martinez, V.; McIntyre, N.E.; Cox, S.; Tissue, D.T.; Zak, J.C. Linking microbial community structure and function to seasonal differences in soil moisture and temperature in a Chihuahuan desert grassland. Microb. Ecol. 2009, 58, 827–842. [Google Scholar] [CrossRef]
- Xiao, C.; Janssens, I.A.; Liu, P.; Zhou, Z.; Sun, O.J. Irrigation and enhanced soil carbon input effects on below-ground carbon cycling in semiarid temperate grasslands. New Phytol. 2007, 174, 835–846. [Google Scholar] [CrossRef]
- Hastings, S.J.; Oechel, W.C.; Muhlia-Melo, A. Diurnal, seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California, Mexico. Glob. Chang. Biol. 2005, 11, 927–939. [Google Scholar] [CrossRef]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in and and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef]
- Xie, J.; Zha, T.; Jia, X.; Qian, D.; Wu, B.; Zhang, Y.; Bourque, C.P.A.; Chen, J.; Sun, G.; Peltola, H. Irregular precipitation events in control of seasonal variations in CO2 exchange in a cold desert-shrub ecosystem in northwest China. J. Arid Environ. 2015, 120, 33–41. [Google Scholar] [CrossRef]
- Turner, M.G.; Dale, V.H. Comparing large, infrequent disturbances: What have we learned? Ecosystems 1998, 1, 493–496. [Google Scholar] [CrossRef]
- Niu, S.; Yang, H.; Zhang, Z.; Wu, M.; Li, L.; Han, X.; Wan, S. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe. Ecosystems 2009, 12, 915–926. [Google Scholar] [CrossRef]
- Kwon, H.; Pendall, E.; Ewers, B.E.; Cleary, M.; Naithani, K. Spring drought regulates summer net ecosystem CO2 exchange in a sagebrush-steppe ecosystem. Agric. For. Meteorol. 2008, 148, 381–391. [Google Scholar] [CrossRef]
- Chen, L.; Li, F.; Di, X.; Zhao, J. Aeolian Sandy Soils in China; Science Press: Beijing, China, 1998. [Google Scholar]
- Sun, Y.; Ding, Y. A projection of future changes in summer precipitation and monsoon in East Asia. Sci. China Earth Sci. 2010, 53, 284–300. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, B.; Lu, Q. Progress in the study on response of arid zones to precipitation change. For. Res. 2012, 25, 100–106. (In Chinese) [Google Scholar]
- Wu, B.; Han, H.; He, J.; Zhang, J.; Cui, L.; Jia, Z.; Yang, W. Field-specific calibration and evaluation of ECH2O EC-5 sensor for sandy soils. Soil Sci. Soc. Am. J. 2014, 78, 70–78. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CO2 Flux | 2016 | 2017 | |
|---|---|---|---|
| Month (M) | GEP | 35.974 *** | 13.709 *** |
| ER | 24.564 *** | 17.487 *** | |
| NEE | 31.505 *** | 10.864 *** | |
| Treatment (T) | GEP | 4.319 * | 6.219 ** |
| ER | 8.756 ** | 15.993 *** | |
| NEE | 3.678 * | 3.106 | |
| Interaction (M × T) | GEP | 1.861 | 1.391 |
| ER | 1.000 | 1.160 | |
| NEE | 2.118 | 1.169 |
| Variable | F | |
|---|---|---|
| Year (Y) | GEP | 5.236 * |
| ER | 55.496 *** | |
| NEE | 0.798 | |
| Treatment (T) | GEP | 24.496 *** |
| ER | 58.812 *** | |
| NEE | 12.773 *** | |
| Interaction (Y × T) | GEP | 1.975 |
| ER | 2.157 | |
| NEE | 2.633 *** |
| Year | Factor | GEP | ER | NEE | |||
|---|---|---|---|---|---|---|---|
| R2 | p Value | R2 | p Value | R2 | p Value | ||
| 2016 | Air temperature | 0.051 | 0.017 | 0.178 | <0.001 | 0.035 | 0.03 |
| Soil temperature | 0.027 | 0.083 | 0.005 | 0.429 | 0.055 | 0.006 | |
| Soil moisture at 10 cm | 0.026 | 0.090 | 0.002 | 0.576 | 0.043 | 0.016 | |
| Soil moisture at 20 cm | 0.000 | 0.848 | 0.033 | 0.040 | 0.129 | <0.001 | |
| 2017 | Air temperature | 0.100 | 0.001 | 0.077 | 0.004 | 0.047 | 0.024 |
| Soil temperature | 0.055 | 0.007 | 0.004 | 0.505 | 0.033 | 0.038 | |
| Soil moisture at 10 cm | 0.004 | 0.486 | 0.103 | <0.001 | 0.009 | 0.275 | |
| Soil moisture at 20 cm | 0.143 | <0.001 | 0.383 | <0.001 | 0.031 | 0.059 | |
| C | A + 50% | A + 100% | |
|---|---|---|---|
| East–west length of the nabkhas (m) | 5.75 ± 0.76 | 6.30 ± 0.89 | 7.00 ± 1.34 |
| South–north length of the nabkhas (m) | 6.45 ± 1.03 | 6.05 ± 0.69 | 8.83 ± 1.83 |
| Height of the nabkhas (m) | 1.25 ± 0.18 | 1.18 ± 0.07 | 1.40 ± 0.23 |
| Area of the nabkhas (m2) | 30.51 ± 7.67 | 29.94 ± 5.06 | 51.64 ± 15.38 |
| Volume of the nabkhas (m3) | 13.98 ± 4.95 | 11.94 ± 2.45 | 26.11 ± 10.20 |
| Plant cover (%) | 25.00 ± 3.54 a | 26.25 ± 5.54 a | 33.75 ± 4.73 a |
| Relative cover of N. t. | 0.74 ± 0.09 a | 0.83 ± 0.06 a | 0.63 ± 0.07 a |
| Canopy height (cm) | 51.88 ± 3.73 a | 60.69 ± 3.69 a | 61.81 ± 2.95 a |
| Soil organic carbon (%) | 0.104 ± 0.012 a | 0.099 ± 0.006 a | 0.157 ± 0.024 b |
| Soil total nitrogen (‰) | 0.143 ± 0.009 a | 0.140 ± 0.004 a | 0.178 ± 0.013 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Wu, B.; Bao, F.; Gao, Y.; Li, X.; Cao, Y.; Lu, Q.; Gao, J.; Xin, Z.; Liu, M. Different Responses of Growing Season Ecosystem CO2 Fluxes to Rain Addition in a Desert Ecosystem. Plants 2023, 12, 1158. https://doi.org/10.3390/plants12051158
Xu X, Wu B, Bao F, Gao Y, Li X, Cao Y, Lu Q, Gao J, Xin Z, Liu M. Different Responses of Growing Season Ecosystem CO2 Fluxes to Rain Addition in a Desert Ecosystem. Plants. 2023; 12(5):1158. https://doi.org/10.3390/plants12051158
Chicago/Turabian StyleXu, Xiaotian, Bo Wu, Fang Bao, Ying Gao, Xinle Li, Yanli Cao, Qi Lu, Junliang Gao, Zhiming Xin, and Minghu Liu. 2023. "Different Responses of Growing Season Ecosystem CO2 Fluxes to Rain Addition in a Desert Ecosystem" Plants 12, no. 5: 1158. https://doi.org/10.3390/plants12051158