Abstract
Numerous environmental conditions negatively affect plant production. Abiotic stresses, such as salinity, drought, temperature, and heavy metals, cause damage at the physiological, biochemical, and molecular level, and limit plant growth, development, and survival. Studies have indicated that small amine compounds, polyamines (PAs), play a key role in plant tolerance to various abiotic stresses. Pharmacological and molecular studies, as well as research using genetic and transgenic approaches, have revealed the favorable effects of PAs on growth, ion homeostasis, water maintenance, photosynthesis, reactive oxygen species (ROS) accumulation, and antioxidant systems in many plant species under abiotic stress. PAs display a multitrack action: regulating the expression of stress response genes and the activity of ion channels; improving the stability of membranes, DNA, and other biomolecules; and interacting with signaling molecules and plant hormones. In recent years the number of reports indicating crosstalk between PAs and phytohormones in plant response to abiotic stresses has increased. Interestingly, some plant hormones, previously known as plant growth regulators, can also participate in plant response to abiotic stresses. Therefore, the main goal of this review is to summarize the most significant results that represent the interactions between PAs and plant hormones, such as abscisic acid, brassinosteroids, ethylene, jasmonates, and gibberellins, in plants under abiotic stress. The future perspectives for research focusing on the crosstalk between PAs and plant hormones were also discussed.
1. Introduction
Several environmental conditions can negatively affect plant production. Abiotic stresses, such as salt, drought, cold, heat, and heavy metal toxicity, cause plant damage at the physiological, biochemical and molecular level [1]. Salinity and drought are significant factors that threaten global agriculture [2,3,4]. Low temperatures are also considered to be a crucial harmful stress [5]. Moreover, elevated concentrations of heavy metals degrade soil health and, consequently, disrupt plant growth [6,7]. Abiotic stresses disrupt the water balance, membrane permeability, mineral nutrients uptake, and accumulation. Furthermore, photosynthesis, respiration, and protein synthesis are also limited. In plants, under various abiotic stresses, an imbalance between the production of reactive oxygen species (ROS) and their metabolism is observed. Oxidative stress negatively affects biomolecules and cellular organelles [8]. In response to unfavorable conditions, plants undergo physiological and metabolic reactions accompanied by modifications at the post-translational and post-transcriptional levels. Enhanced polyamines (PAs) accumulation is one of the most remarkable metabolic signs in plants subjected to abiotic stressors [5]. PAs are ubiquitous, polycationic, and aliphatic amines that play a significant role in physiological and developmental processes such as organogenesis, embryogenesis, cell division, floral development, reproductive organ development, root growth, leaf senescence, and fruit maturation [9,10]. PAs are also involved in plant response to biotic and abiotic stresses such as salinity, drought, heavy metals, and high- and low-temperature stresses [11,12,13,14]. Studies have shown that PAs display multi-track actions in plant response and stress tolerance. PAs improve the stability of membranes, DNA, and other bioactive molecules. They participate in chromatin condensation, maintenance of DNA structure, RNA processing, translation, and protein activation [15,16,17]. During abiotic stress, PAs modulate the expression of genes related to starch metabolism, photosynthesis, antioxidant machinery, and stress response [18,19,20,21]. PAs also play a significant role in the regulation of ion uptake and maintenance of ionic homeostasis [22]. For example, PAs directly inhibit fast- and slow-acting non-selective cation channels (NSCCs) in tonoplast with high affinity and reduce the efflux of toxic Na+ ions from vacuole to cytoplasm during salinity [23,24,25]. PAs can also activate Ca2+-ATPase localized in the plasma membrane and tonoplast, thereby regulating Ca2+ transport and preventing excessive Ca2+ level during prolonged exposure to stress [25,26,27]. PAs positively affect antioxidant machinery, improve osmolyte accumulation, enhance photosynthesis, and maintain nitrogen metabolism [13,28,29,30]. PAs interact with plant hormones, such as abscisic acid (ABA), ethylene (ETH), jasmonates (JAs), and gibberellins (GAs), and signaling molecules, such as γ-aminobutyric acid (GABA) and nitric oxide (NO), to enhance plant tolerance of abiotic stresses [31,32,33,34,35]. They are also involved in the production of a signal molecule, hydrogen peroxide (H2O2), due to PAs catabolism [36]. A consequence of PA-induced H2O2 generation may be the induction of stress-response genes or programmed cell death (PCD) [37,38] (Figure 1).
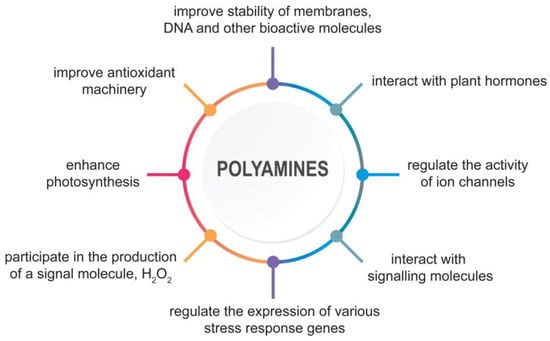
Figure 1.
The multitrack action of PAs in plant response and tolerance to abiotic stresses.
Plant hormones, commonly known as plant growth regulators, control growth and developmental processes that play a meaningful role in plant adaptation to unfavorable environments. They are divided into two groups: (I) positive growth regulators, such as auxins, cytokinins (CKs), GAs, and brassinosteroids (BRs); and (II) stress hormones, such as ABA, JAs, salicylic acid (SA), and ETH [39]. The additive effect of phytohormones on plant tolerance of abiotic stresses has been known for many years and has been extensively reviewed [40,41,42,43]. The literature has shown that plant hormones interact with PAs during plant growth and development (Table 1). Plant hormones regulate the expression of PAs metabolic genes [44,45,46]. On the other hand, PAs can regulate the biosynthesis, signal transduction, and transport of certain plant hormones [47,48,49]. Interestingly, some studies have revealed that plant hormones can participate in plant defense through interactions with PAs, inducing plant tolerance of abiotic stresses [39,50,51]. Although it has been known that PAs can interact with plant hormones, recent studies have gradually brought us closer to understanding the mechanisms of crosstalk between PAs and phytohormones during abiotic stress. Therefore, this paper aims to summarize the data on the relationship between PAs and hormones in plants subjected to various abiotic stresses. The potential direction for further studies related to this research area, which may lead to the discovery of detailed molecular mechanisms, is discussed.

Table 1.
Representative evidence for the interaction between PAs and plant hormones during plant growth and development. 6-BA, 6-benzylaminopurine; IAA, indole acetic acid; iP, isopentenyl adenine; NaSA, sodium salicylate.
2. PAs Metabolism
The most common PAs in the plant kingdom are diamine putrescine (Put), triamine spermidine (Spd) and tetraamine spermine (Spm) [12]. Other PAs identified in plants include agmatine (Agm), cadaverine (Cad), and an isomer of Spm, thermospermine (tSpm) [5,34]. The routes of PAs biosynthesis and catabolism, as well as PAs back-conversion, are shown in Figure 2.
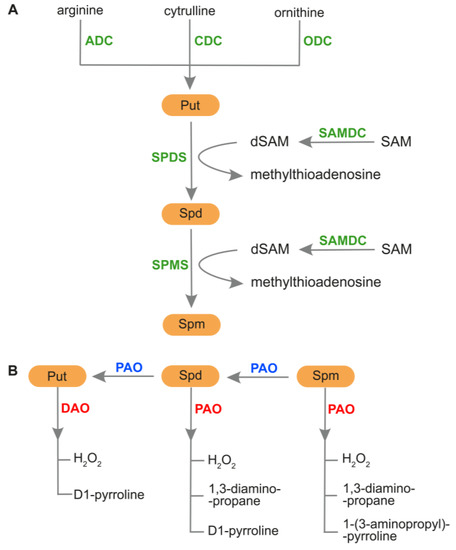
Figure 2.
Metabolism of PAs in plants. (A) PAs biosynthesis. Production of Put can be catalyzed by ADC form arginine, by ODC from ornithine, or by CDC from citrulline. In the next step, SPDS is responsible for biosynthesis of Spd from Put, and SPMS participates in the production of Spm from Spd. During this process, dSAM is used as an aminopropyl donor to produce Spd and Spm. An enzyme, SAMDC takes part in the conversion of SAM to the form of dSAM. (B) PAs catabolism and back-conversion. DAO catalyzes Put oxidation during which D1-pyrroline and H2O2 are generated. PAO plays a role in Spd and Spm oxidation, producing H2O2, 1,3-diaminopropane and 1-pyrroline or 1-(3-aminopropyl)-pyrroline. PAO can also participate in the back-conversion of PAs, during which Spm is converted into Spd and next into Put. ADC, arginine decarboxylase; CDC, citrulline decarboxylase; DAO, diamine oxidase; dSAM, decarboxylated S-adenosylmethionine; ODC, ornithine decarboxylase; PAO, polyamine oxidase; Put, putrescine; SAM, S-adenosylmethionine; SAMDC, S-adenosylmethionine decarboxylase; Spd, spermidine; SPDS, spermidine synthase; Spm, spermine; SPMS, spermine synthase.
The production of Put in plant tissues is catalyzed by arginine decarboxylase (ADC) or ornithine decarboxylase (ODC). However, some plants, such as Arabidopsis thaliana and many members of the Brassicaceae family, have lost their ODC genes [35,66]. Another pathway for Put production has been reported in sesame, in which Put biosynthesis occurs from citrulline (Cit) in a reaction catalyzed by citrulline decarboxylase (CDC) [66]. After Put production, S-adenosyl-L-methionine decarboxylase (SAMDC) introduces S-adenosyl-L-methionine (SAM) into the pathway. Then, the decarboxylated form of SAM (dSAM) is used as an aminopropyl donor in the conversion of Put to Spd and then Spd to Spm. The reaction is catalyzed by Spd synthase (SPDS) and Spm synthase (SPMS), respectively [66]. Two enzymes, diamine oxidase (DAO; also known as CuAO) and polyamine oxidase (PAO), are involved in PAs catabolism. DAO catalyzes Put oxidation in the apoplast, resulting in the production of D1-pyrroline and H2O2. PAO participates in Spd and Spm oxidation in apoplast and peroxisomes, generating H2O2, 1,3-diaminopropane and 1-pyrroline or 1-(3-aminopropyl)-pyrroline [64,67,68,69,70]. The product of PAs catabolism, H2O2, plays a significant role in plant development and response to environmental stresses; however, overaccumulation of this molecule negatively affects plants [14,51,69]. It is worth noting that other products of PA catabolism are acrolein and GABA [14]. Some PAOs catalyze the back-conversion of higher PAs. During this process, Spm is converted into Spd and subsequently into Put [71].
3. Interaction of PAs and Plant Hormones under Abiotic Stress
The literature has indicated that PAs play a significant role in plant tolerance of abiotic stresses [5,64]. Furthermore, many studies have revealed that plant hormones are important elements in plant responses to adverse environmental conditions [41,70]. These two groups of molecules appear to be significant in the signal transduction pathway that is activated in plants in response to abiotic stress. Determining the interactions between PAs and plant hormones is crucial. Therefore, reports indicating crosstalk between PAs and phytohormones and their significance in improving plant tolerance to abiotic stresses have been summarized (Table 2).
Table 2.
Summary of evidence for the interaction between PAs and plant hormones in response to abiotic stresses.
3.1. PAs-Plant Hormones Crosstalk under Drought Stress
Drought is considered to be one of the most threatening abiotic stresses in plants. It has a negative effect on plant growth and crop productivity [39]. Drought leads to the loss of cell turgor, water and mineral disruption, and photosynthesis limitation [3,83]. A negative effect of drought is osmotic stress, which induces oxidative stress. Excessive accumulation of ROS can disrupt the functioning of cell membranes and induce lipid peroxidation and DNA modifications. This leads to metabolic and structural dysfunction and PCD [13,79].
The essential mechanisms of plant response to drought include the participation of ABA. This phytohormone is involved in seed dormancy, embryo morphogenesis, and fat and stored protein production [43]. During these processes, ABA interacts with multiple plant hormones, signaling molecules, and secondary metabolites such as benzoxazinoids [72,94,95,96]. During drought, ABA stimulates stomatal closure to prevent excessive water loss from leaves, modulates the expression of multiple genes involved in the plant response to stress, and modifies the biosynthesis of osmoprotectants [21]. Studies have revealed that exogenous ABA increased plant tolerance to drought by improving cell membrane stability, stimulation of antioxidant enzymes, and reduction of ROS content [43]. Previous studies have provided evidence for crosstalk between PAs and ABA (Figure 3A). Exogenous PAs induce stomatal closure in Vicia faba guard cells [73]. On the other hand, ABA has been reported to stimulate the activity of CuAO and NADPH oxidase, leading to enhanced H2O2 production in guard cells. Consequently, H2O2 induces Ca2+ influx into guard cells, promoting stomatal closure [73]. Comprehensive studies of Arabidopsis thaliana, have also revealed the role of PAs in regulating stomatal movement. It has been suggested that Spm may contribute to increasing cytoplasmic Ca2+ concentration by activating specific ion channels, especially Ca2+-permeable channels, which inactivate the K+ inward rectifier at the plasma membrane. This could stimulate stomatal closure [11]. Furthermore, the application of exogenous PAs increase NO and ROS levels in guard cells. PAs induce ROS production via NADPH oxidase and amine oxidase (AO) [75]. It has been suggested that the action of PAs and ABA is linked during stomatal closure due to the involvement of the same key components: ROS and NO [75,97]. The relationship between PAs and ABA in the regulation of stomatal movement in Arabidopsis is supported by Fincato et al. [74]. The ABA-induced expression of AtPAO2 has also been reported in guard cells. Moreover, Fraudentali et al. [98] described that AtCuAOδ participates in ABA-induced stomatal closure.
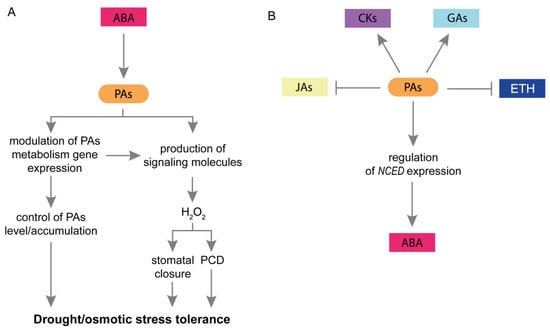
Figure 3.
Interaction between PAs and plant hormones in response to drought and osmotic stress. (A) The effect of ABA on PAs’ mode of action. ABA modulates the expression of PAs metabolism genes and controls PAs content, thereby inducing plant tolerance to drought and osmotic stress. Furthermore, due to its impact on PAs catabolism, ABA regulates the production of H2O2, which may induce stomatal closure or PCD. (B) A schematic representation of possible PA-plant hormone interactions during plant responses to drought and osmotic stress. During drought/osmotic stress, PAs regulate NCED expression and ABA content. They also increase CKs and GAs content and reduce ETH and JAs content, probably inducing plant tolerance to stress. ABA, abscisic acid; CKs, cytokinins; ETH, ethylene; GAs, gibberellins; H2O2, hydrogen peroxide; JAs, jasmonates; NCED, 9-cis-epoxycarotenoid dioxygenase; PAs, polyamines; PCD, programmed cell death.
It has also been proposed that PAs are involved in plant tolerance and acclimation to drought by modulating ABA biosynthesis. Appropriate ABA content is important for plant stress tolerance. Excessive ABA accumulation may lead to reduced plant tolerance to drought [83]. Marcińska et al. [82] examined the effect of exogenous PAs on stress tolerance in two wheat genotypes exposed to osmotic stress. In drought-resistant plants, exogenous PAs (especially Spd) reduce the ABA content during osmotic stress. This may indicate the role of PAs in alleviating the adverse effects of stress and the negative physiological effects of drought. In contrast, in drought-sensitive wheat seedlings, the application of PAs (especially Spm) led to further increases in the ABA content. The authors suggested that an increase in ABA levels reflects a high intensity of drought response. Plants attempt to avoid the lethal effects of stress and reduce water loss through stomata [82]. PAs may regulate ABA content at the genetic level by modulation of genes encoding ABA biosynthesis enzymes. ABA is produced from C40 carotenoids in three steps catalyzed by zeaxanthin epoxidase, 9-cis-epoxycarotenoid dioxygenase (NCED), and abscisic aldehyde oxidase (AAO) [99,100,101]. Put strongly enhanced the expression of NCED in wheat leaves under osmotic and drought conditions [78,79].
Several studies have described the relationship between ABA and PAs metabolism. Exogenous ABA up-regulates the expression of several genes involved in PA production in Arabidopsis [45]. PAO activity and expression of PAO were induced by exogenous ABA [52,54]. A complex expression profile of seven PAO genes was also presented in tomato plants treated with exogenous ABA [44]. During drought, ABA may induce synchronized effects on the anabolism/catabolism of PA to increase the cellular ROS load for signaling downstream stress defense events. In Vitis vinifera, ABA stimulates the activities of enzymes involved in PA biosynthesis and, at least partially, induces PA accumulation and their exodus into the apoplast. At the same time, ABA is an upstream signal for the induction of PAs catabolism. Accumulated PAs begin to oxidize primarily in the apoplast. A product of PAs oxidation, H2O2, may act as a secondary messenger in the signaling pathway, inducing stomatal closure and/or PCD [102,103]. Furthermore, ABA has been shown to modulate the expression of PAs production and catabolism genes in response to drought [81,104].
Higher ABA and PAs contents were observed under drought conditions in the transgenic Arabidopsis line overexpressing OsHSFA3, which encodes one of the heat-shock factors associated with improved tolerance to drought and high temperature [79]. This was related to the higher expression of the PAs biosynthesis genes (ADC1, ADC2, SPDS1, and SPMS). Furthermore, ABA-responsive elements (ABREs) have been reported in the promoter regions of these PAs biosynthetic genes, suggesting a role in the regulation of PA production [76,77]. Moreover, in the Arabidopsis exposed to drought, the expression of ABA-inducible and drought-responsive genes (RD29A and RD22) increased and showed similar kinetics as the expression of PAs biosynthesis genes (ADC1, ADC2, SPDS1, SPDS2, SPMS, and SAMDC1). This supports the role of ABA signaling in the regulation of PAs production at the transcriptional level [68]. In silico analysis by Basu et al. [78] revealed the presence of several putative cis-acting elements, such as ABRE, Low-Temperature Responsive Element (LTRE), MYB, and W-box, in the promoter region of rice SAMDC. These elements are strongly associated with environmental factors such as drought, cold, and ABA signaling [78].
Several studies have suggested a relationship between PAs and other phytohormones in plant response to drought (Figure 3B). For example, the application of exogenous PAs modulated GA and SA content in some plant species under drought conditions [39,83,85,105], probably increasing plant tolerance to stress. However, the physiological implications of these changes require further investigation. BRs are steroidal plant hormones that regulate plant growth and productivity. The exogenous application of BRs has revealed their role in plant tolerance to low and high temperatures, heavy metals, drought, salinity, and waterlogging [13,106]. Talaat et al. [13] reported that the co-application of 24-epibrassinolide (EBL), one of BRs forms, and Spm improved plant drought tolerance and reduced the accumulation of ROS by enhancing their scavenging through the elevation of antioxidant machinery. In this case, further studies are necessary to reveal mechanisms between PAs and BRs in plant response to this stress.
JAs are other plant growth regulators involved in plant responses to stress through crosstalk with PAs. The JAs family includes methyl jasmonate (MeJA), jasmonic acid (JA), and jasmonyl-isoleucine [107]. JAs regulate the expression of genes responsible for tolerance, interact with other phytohormones, and modulate proteome profiles. They can function as stress modulators by suppressing or enhancing plant stress responses [108]. Peremarti et al. [84] showed that in rice MeJA negatively regulated OsAdc, OsSamdc, and OsSpds gene expression. According to the obtained results, it was proposed that drought-induced Put accumulation occurs until the threshold for conversion to higher PAs is reached. At the same time, activation of JAs signaling is important for the enhancement of Put conjugation, reduction of free Put content, and withdrawal of plants from the threshold [84]. On the other hand, PAs decrease JAs accumulation in plants exposed to drought [43]. It is well known that the application of JAs retards plant growth, and activation of JAs signaling causes repression of photosynthesis and photosynthesis-related gene expression [109,110]. Therefore, it is suggested that PA-mediated reduction of JAs content may play a vital role in plant tolerance to stress [39].
More advanced studies have presented the crosstalk between PAs and ETH at several levels. ETH is a gaseous plant hormone that modulates a broad array of plant responses such as cell expansion, seed germination, flowering, fruit ripening, and senescence. It also affects growth parameters, osmolytes and pigment content, photosynthesis, and oxidative stress in plants subjected to different abiotic stresses [111]. It is suggested that ETH participates in the regulation of stomatal movement. However, the detailed role of this plant hormone in plant responses to drought has not been well established [43]. Interestingly, PAs and ETH are connected in their biosynthetic pathways. They have the same precursor, SAM, produced from L-methionine and ATP by S-adenosyl-L-methionine synthetase (SAMS) [33]. Hence, it has been suggested that PA and ETH compete for SAM production [31]. Recent studies have demonstrated that PAs regulate ETH content in some plant species. Grzesiak et al. [112] suggested that higher PAs content led to a reduction of ETH biosynthesis in PEG-treated wheat seedlings. Liu et al. [83] showed that the effect on ETH content might depend on the PAs’ type and lead to different results. A reduction in the ETH evolution rate was observed in the grain-filling of wheat under drought conditions. Application of Spd and Spm during drought resulted in further reduction of ETH evolution rate, whereas CKs and ABA content increased, thereby promoting wheat grain filling under drought conditions. Interestingly, an adverse effect was observed for exogenous Put, which increased ETH evolution rate and led to excessive ABA accumulation and inhibition of wheat grain filling [83]. Furthermore, Huang et al. [3] demonstrated that the activity of ribulose 1,5-bisphosphate carboxylase (RuBPC) was reduced in plants in response to drought. Aminooxyacetic acid (AOAA, an inhibitor of ethylene synthesis) may enhance RuBPC activity by modulating PAs levels and decreasing the damage caused by ETH release [3]. Additionally, a close relationship between PAs, ETH, and ROS was observed in the leaves of Glycyrrhiza inflata seedlings under osmotic stress. In severely damaged leaves, ROS production was modulated by PAs and ETH. Inhibition of ETH generation decreased ROS production, whereas a reduction in PAs content promoted ROS generation. Conversely, the exogenous application of H2O2 promoted ETH production and reduced PAs content [31].
3.2. PAs-Phytohormones Cross Talk under Salt Stress
Salinity is another factor that can reduce agricultural productivity [85]. According to estimates, salinity affects more than 800 million hectares of arable land worldwide. Salt stress reduces plant growth and disrupts photosynthesis and respiration [85]. Salinity causes the overproduction and accumulation of ROS in plants, which leads to salinity-induced osmotic stress [113,114].
Among all plant hormones, crosstalk between PAs and ETH during salt stress seems to be the most established. It has been suggested that plant tolerance to salinity may be related to ETH biosynthesis and signal transduction pathways. Exogenous application of ETH increased plant tolerance to salinity due to the increased expression of genes involved in ROS scavenging [115]. Furthermore, ETH may help regulate Na+ and K+ concentrations in xylem tissues under salt stress [111]. In contrast, the application of ETH biosynthesis inhibitors increases plant sensitivity to salt stress [115]. PAs can play a role in modulating ETH production during salinity. Quinet et al. [67] demonstrated that the application of exogenous Put enhanced ETH synthesis in the salt-resistant cultivar exposed to salinity [67]. Zapata et al. [115] noticed that under salinity, the ETH, Spd, and Spm contents increased in plant species with different salt sensitivity. PAs and ETH may compete for SAM production. However, the results support that no competence between them occurred, and the SAM pool is high enough to support both PAs and ETH biosynthesis during salt stress [31,115]. Because PAs catabolism is important in plant responses to stress, the crosstalk between PAs catabolism and ETH signaling has been investigated. In transgenic tobacco plants displaying reduced PAO activity, the transcript levels of genes encoding 1-amino-cyclopropane-carboxylate synthase (ACS) and 1-amino-cyclopropane-carboxylate oxidase (ACO) were higher than those in plants displaying enhanced PAO activity [4]. ACS converts SAM to 1-aminocyclopropane-1-carboxylic acid (ACC), whereas ACO converts ACC to ETH [116]. To provide further insight into the link between PAs, ETH, and H2O2, Freitas et al. [70] proposed a detailed model of salinity-induced expression and activity of ACO, leading to enhanced ETH biosynthesis in the salt-tolerant maize genotype. Consequently, ETH induces H2O2 production due to enhanced DAO expression and activity. H2O2 plays a dual role. It acts as a signaling molecule for PAs synthesis during the first stage of stress. In the second stage of stress, H2O2 inhibits “stress” ETH production and activates PAs biosynthesis by up-regulating ADC expression and activity. A recent study provided by Takács et al. [46] demonstrated that ETH-mediated signaling associated with NR receptors played a key role in PA metabolism in tomato fruits at the mature stage during salt stress conditions. In this study, the tomato ripening mutant Never ripe (Nr), which has a mutation in the ETH-binding domain of the ETH receptor NR, was used [117]. Among the three PAs tested, salt stress enhanced only Spm content in WT fruits. However, this change was not observed in Nr fruits exposed to salt stress. Moreover, during salinity stress, the activities of DAO and PAO were strongly stimulated in Nr fruits, contributing to higher oxidative stress [46].
BRs positively influence plant tolerance to salt stress. In Arabidopsis, enhanced BR signaling activity increases salt stress tolerance. In contrast, the BR-defective mutants exhibit hypersensitivity to salinity. Moreover, BRs may control water loss by reducing stomatal density in plants exposed to high salt stress conditions [40]. Co-application of BRs and PAs has a beneficial effect on plants exposed to salt stress [106,118]. Exogenous application of BRs enhances plant tolerance to salinity by modulating PAs homeostasis and distribution in plants [59,86]. Liu et al. [14] described a more advanced model of PAs and BRs interactions during plant response to low and high salt salinity conditions. EBL promotes tolerance of canola to high salt salinity, but not to low salt stress. The dual effect of EBL on plant responses to low and high salinity is related to H2O2 accumulation, which is regulated by PAs metabolism (especially Put oxidation). Under high-salt stress, EBL reduced the accumulation of H2O2 and DAO activity. Interestingly, EBL modulated these parameters in the opposite manner in seedlings exposed to low-salt stress [14].
A high endogenous ABA content is important for increasing plant tolerance to salt stress. ABA induces stomatal closure and accumulation of numerous proteins and osmoprotectants [40]. In the atpao5-3 mutant, salt stress increased ABA level, up-regulated NCED expression and enhanced ABA-inducible RD29B expression [64]. On the other hand, the expression of NCED3 was reduced in Spm-deficient spms mutant and double acl5/spms mutant under salt stress conditions. In addition, in transgenic Arabidopsis lines overexpressing SAMDC1, two of the five ABA-induced genes (NCED and RD29A) were up-regulated by salinity [81] (Figure 4B).
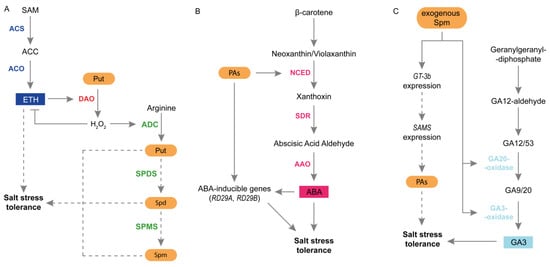
Figure 4.
Interactions between PAs and plant hormones in plants subjected to salt stress. (A) Crosstalk between PAs and ETH during salinity stress in maize. In maize exposed to salt stress, the expression of ACS and ACO increased, leading to a higher ETH content. As a result, ETH stimulates DAO expression and activity, which oxidizes Put to H2O2. Next, H2O2 plays a dual role. It reduces ETH production and prevents excessive ETH content, which may have an unfavorable effect on plants. Moreover, H2O2 stimulates PA production by upregulating ADC expression and ACD activity. (B) Interactions between PAs and ABA in plant response to salt stress. PAs regulate the expression of NCED and ABA content. PAs also regulate the expression of other ABA-inducible genes, such as RD29A and RD29B, thereby inducing plant tolerance to salt stress. (C) PAs and GAs relationships during plant response to salt stress. Exogenous Spd stimulates the activity of GA20 oxidase and GA3 oxidase and up-regulates the expression of some genes involved in GA biosynthesis. Under salt-stress conditions, exogenous Spd enhances GA3 content and stimulates expression of the GT-3b transcription factor. Furthermore, exogenous Spd may promote the expression of SAMS via GT-3b and PAs production, thereby increasing plant tolerance to salt stress. AAO, abscisic aldehyde oxidase; ACC, 1-aminocyclopropane-1-carboxylic acid; ACO, 1-amino-cyclopropane-carboxylate oxidase; ACS, 1-amino-cyclopropane-carboxylate synthase; ADC, arginine decarboxylase; DAO, diamine oxidase; ETH, ethylene; GA, gibberellin; H2O2, hydrogen peroxide; NCED, 9-cis-epoxycarotenoid dioxygenase; PAs, polyamines; Put, putrescine; SAM, S-adenosyl-L-metionine; SAMS, S-adenosyl-L-metionine synthase; SDR, short-chain alcohol dehydrogenase/reductase; Spd, spermidine; SPDS, spermidine synthase; Spm, spermine; SPMS, spermine synthase.
GAs can also interact with PAs during plant response to salt stress. GAs are tetracyclic diterpenoid carboxylic acids that participate in processes related to plant growth and development [119]. GAs positively regulate germination, leaf expansion, and stem elongation. They also initiate flowering, trichome formation, and reproductive development [1]. Salt stress negatively affects endogenous GAs content in plants. This was accompanied by a higher accumulation of DELLA proteins, which are major GA-negative regulators. Interestingly, studies have shown that GA-deficient biosynthetic or signaling mutants exhibit tolerance to severe salt stress. In contrast, the quadruple DELLA loss-of-function mutant was less tolerant to salt [40]. Recent studies have shown that exogenous GAs improve plant tolerance to salinity. Positive effects of Spd and GA3 on priming-induced physiological and biochemical changes have been reported in salt-tolerant and salt-sensitive rice cultivars [2]. In cucumbers, exogenous Spd up-regulated the expression of genes related to GA biosynthesis and increased the activity of GA3-oxidase and GA20-oxidase. Moreover, in salt-stressed plants, exogenous Spd stimulated the accumulation of GA3 and enhanced the expression of the GT-3b transcription factor, a stress-related protein usually induced by salt stress. Spd-induced salt tolerance was reduced in plants treated with GA biosynthesis inhibitor. This suggests that GA plays a role in Spd-induced salt tolerance in cucumber [33] (Figure 4C). In the same species, the application of Spd to salt-treated plants reduced JA content [87]. The role of PAs in the regulation of JA content has also been demonstrated in Arabidopsis seedlings exposed to salinity. Exposure of the atpao5-3 mutant to salt stress resulted in strongly enhanced JA accumulation [64].
The relationship between CKs and PAs in plant responses to salt stress has not been well described. CKs such as zeatin (Z) and zeatin ribose (ZR) are involved in plant cell division and growth, morphogenesis, chloroplast biogenesis, nutrient absorption and balance, and vascular differentiation. They also participate in seed germination and delay senescence in plants [39,51,83]. CKs play an essential role in plant responses to nutrient, drought, salinity, and temperature stresses [120]. Simultaneous application of kinetin (KN) and PAs improves tolerance to salt stress in some plant species, such as Vigna sinensis, wheat, and Luffa acutangular [50,51,88]. The positive influence of KN and Spm on photosynthesis parameters has been extensively described in wheat exposed to salt stress. Grain priming with KN, Spm, or KN + Spm alleviated the negative effects of stress by stimulating leaf area expansion, pigment production, photosynthetic activity, and improving chloroplasts ultrastructure [50]. Additionally, Kapoor and Hasanuzzaman [51] reported that exogenous KN and Put synergistically mitigate salt stress in sponge gourd (Luffa acutangula L.). Simultaneous application of KN and Put to salt-stressed seedlings resulted in a significant enhancement of growth, photosynthetic pigment content, and osmolyte accumulation. Moreover, the co-application of KN and Put up-regulated the antioxidant enzymes activity and increased the content of the non-enzymatic components of the antioxidant defense machinery. Simultaneous KN and Put treatment was more effective than individual application of KN or Put, suggesting the presence of regulatory crosstalk mechanisms between KN and Put [51]. However, the more advanced relationship between PAs and CKs remains unknown.
3.3. PAs-Phytohormones Cross Talk under Heavy Metal Stress
Micronutrients such as copper (Cu), iron (Fe), manganese (Mn), nickel (Ni), and zinc (Zn) are heavy metals. These elements are significant in plants, but high concentrations of them have adverse effects [121]. In addition, other heavy metals, such as cadmium (Cd) and lead (Pb), present in the soil can cause stress conditions in plants. Uncontrolled accumulation of heavy metals in the soil has a negative impact on agriculture. Heavy metals disrupt plant growth, senescence, photosynthetic machinery efficiency, and enzymatic activity [89,90].
Some reports suggest that PAs may interact with phytohormones, especially BRs and ETH, during plant responses to heavy metal stress [6,28] (Figure 5). However, the detailed mechanism of this cooperation has not yet been elucidated. The application of BRs and PAs enhanced the tolerance of some plant species to stress caused by Cr (VI) [28,56], Cu (II) [89,122], Mn [121], and Pb [90] (Figure 5A). Simultaneous BRs and PAs treatment improved growth parameters, reduced malondialdehyde (MDA) and H2O2 levels, and enhanced the antioxidant system [28,90,121,122]. To prevent heavy metal ion toxicity, plants have developed strategies to manage ion uptake, translocation, and distribution, involving BRs and PAs [42,122]. In plants exposed to Cu (II) stress, exogenous Spd and BRs down-regulated the expression of COPT1 and COPT2, which encode the plasma membrane transporters responsible for Cu uptake into the cells. Furthermore, the co-application of Spd and BRs reduced the expression of the HMA5 gene encoding heavy metal ATPases (HMAs), which play an important role in the transmembrane transport of Cu. Finally, the application of Spd and BRs up-regulated the expression of MT1C and CCH1, which encode the proteins involved in Cu detoxification [122]. Heavy metal sequestration is also a crucial process in plant responses to heavy metal stress. Biomolecules such as GSH and phytochelatins can act as metal-chelating agents [42]. Co-application of BRs and PAs increased phytochelatin and GSH content, as well as photosynthetic pigment content [28,122]. Moreover, EBL changed PAs accumulation in mustard under Cu (II) stress. The authors explained that the higher PA content (Spm, Spd, Put, and Cad) in the leaves of mustard exposed to Cu helped the plants mitigate this stress [89].
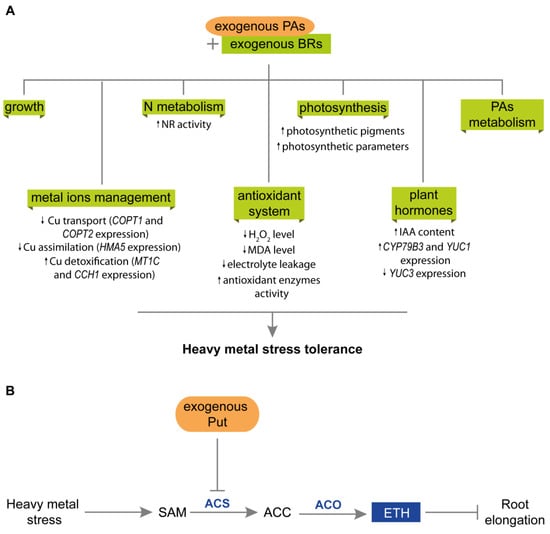
Figure 5.
Interactions between PAs and plant hormones during plant responses to heavy metal stress. (A) Synergistic action of exogenous PAs and BRs in plants exposed to heavy-metal stress. PAs and BRs possibly cooperate at several levels, improving growth parameters, N metabolism, photosynthesis efficiency, and regulating PAs metabolism. They also manage metal ions uptake, distribution, and detoxification and enhance antioxidant machinery, thereby reducing the damage caused by ROS. The co-action of PAs and BRs leads to the enhanced expression of IAA metabolic genes and higher IAA content, probably stimulating plant growth and tolerance to stress. (B) Interactions between PAs and ETH during root elongation in plants exposed to heavy-metal stress. During heavy metal stress, ETH production increases, leading to inhibition of root elongation in plants. However, exogenous Put reduces ACS activity and ETH production; therefore, the inhibition of root elongation is declined. ACC, 1-aminocyclopropane-1-carboxylic acid; ACO, 1-amino-cyclopropane-carboxylate oxidase; ACS, 1-amino-cyclopropane-carboxylate synthase; BRs, brassinosteroids; ETH, ethylene; IAA, indole acetic acid; PAs, polyamines SAM, S-adenosyl-L-metionine.
During heavy metal exposure, ETH production increases in some plant species [6]. ETH signaling is involved in the response of plants to oxidative stress induced by heavy metals. The exogenous application of ETH or ethephone mitigated the negative effects of heavy metal stress due to glutathione accumulation. Additionally, overexpression of ERF genes lead to increased expression of glutathione synthesis genes and increased Cd tolerance [42,111]. One of the adverse effects of plant exposure to Al toxicity is inhibition of root elongation. PAs and ETH are involved in this process [6] (Figure 5B). In wheat cultivars exposed to Al toxicity, Put supplementation alleviates the negative effects of stress. Further analyses revealed that ACS- and ACO-mediated Al-induced ETH production may play a role in Al-induced inhibition of root elongation. Application of Put decreases ACS activity and ETH production and reduces the inhibition of root elongation in plants exposed to Al toxicity [6]. Kohli et al. [90] noted the enhancement in Spd level in roots of mustard treated with EBL + SA and exposed to Pb stress. The positive effect of Spd on plant tolerance may be related to its antioxidant properties, which protect DNA from oxidative damage [90]. Shah et al. [7] described the positive correlation between PAs and ETH in cucumber seedlings supplemented with 2-hydroxymelatonin (2-OHMT) and exposed to Cd stress. In stressed seedlings, the application of 2-OHMT reduced PAO activity, leading to higher Put, Spd, and Spm content. The application of 2-OHMT also enhanced the expression of a gene encoding the ETH receptor (CS-ERS) during stress conditions [7].
3.4. PAs-Phytohormones Cross Talk under Low Temperature Stress
Low temperatures affect many physiological processes such as photosynthesis, nutrient absorption and metabolism, and can cause metabolic disruption and structural damage [5,35,123].
PAs probably maintain the proper ABA content in plants exposed to low-temperature stress. Higher ABA content enhanced tolerance of chilling by inducing stomatal closure and promoting the expression of cold-induced genes [91]. In an Arabidopsis subjected to low temperatures, a rapid increase in Put level is necessary for ABA accumulation [12]. Furthermore, Put enhances NCED expression in stressed plants [12]. In tomato plants exposed to chilling, exogenous Put increased ABA content and triggered the expression of LeNCED1. This indicates that ABA is involved in Put-induced tolerance to chilling stress [35]. Furthermore, ADC2 contains an ABRE transcription factor in its’ promoter. Under low temperature stress, ABRE is responsible for the enhanced expression of genes mediated by ABA [77,124]. Li et al. [123] confirmed the role of CmADC as a positive regulator of melon-seedling cold tolerance and found two transcription factors, ABRE-binding factor 1 (CmABF1), and C-repeat binding factor 4 (CmCBF4), that directly target CmADC to enhance its expression.
The crosstalk between PAs and JAs is also important for plant tolerance to low-temperature stress. In the exocarp of zucchini squash fruit (Cucurbita pepo L.) exposed to chilling, MeJA decreased Put accumulation, whereas Spd and Spm accumulation increased [92]. In mangoes and apples, higher free Spd and Spm content correlated with low-temperature stress tolerance [125,126]. In rice treated with MeJA before chilling, a significant increase in free Put and Spm accumulation in the shoots and roots was observed. In contrast, the free Spd content was reduced. Further analyses have revealed that in rice seedlings, accumulation of Put induced by MaJA is the result of the activation of the ADC pathway, but not the ODC pathway [32]. In Indica-japonica hybrid rice, exogenous Spd enhanced CK and GA3 content during chilling, indicating a possible crosstalk between PAs and these plant hormones [93].
4. Conclusions and Future Perspectives
Permanent changes in environmental conditions challenge equitable agriculture and efficient plant production. The increasing mean air temperature and the accompanying drought, as well as soil salinity, are environmental factors that are subject to rapid changes in the coming years. The unfavorable direction of these changes affects plant growth and development. Therefore, one of the priorities of the scientific world is to acquire and enhance knowledge about the mechanisms that allow plants to deal with abiotic stresses and to develop strategies to introduce transgenic plants to agriculture. Exploration of PAs’ mode of action in plant tolerance of abiotic stress seems to be a crucial step in this process. An important aspect of PAs’ action in the response to abiotic stress is their interactions with plant hormones.
This review presents the relationship between PAs and plant hormones in plant tolerance of abiotic stress (Table 2). Our study has shown the favorable effect of the co-application of exogenous PAs and phytohormones, such as CKs and BRs, on stressed plants. We have demonstrated that PAs affect the accumulation of plant hormones and the expression of their biosynthesis genes during abiotic stress. This study is a good starting point for further research focusing on the regulation of PAs and plant hormones metabolism at the post-transcriptional and post-translational levels in plants under abiotic stress. Furthermore, it is worth investigating the possible function of PAs in the regulation of plant hormone signaling during abiotic stress. Several components, such as Ethylene Response Sensor 1 (ERS1), Ethylene Insensitive 2 (EIN2), EIN3-like (EIL), and Ethylene Responsive Factors (ERF), participate in ETH signaling and play a relevant role in plant response to salt stress [127,128,129]. PAs regulate some components of the ETH signaling pathway [63]. It would be interesting to explore the connections between PAs and plant hormones—such as ETH, ABA, GAs, and JAs signaling pathways during abiotic stress—and establish the role of those relationships in inducing plant tolerance of adverse environmental conditions.
In addition, it seems interesting to examine the role of NO in this aspect. Similar to PAs, NO interacts with plant hormones during plant responses to abiotic stress [130]. Therefore, it would be interesting to investigate the possible mechanisms involving PAs, NO, and plant hormones that induce plant tolerance to stress. A crucial step in achieving this goal is to establish the role of PAs in NO production.
Author Contributions
Conceptualization—N.N.; writing—original draft preparation, N.N.; writing—review and editing, N.N., M.R. and M.J.; visualization, N.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors report no declaration of interest.
References
- Banerjee, A.; Roychoudhury, A. The Regulatory Signaling of Gibberellin Metabolism and Its Crosstalk With Phytohormones in Response to Plant Abiotic Stresses. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 333–339. ISBN 978-0-12-816451-8. [Google Scholar]
- Chunthaburee, S.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Alleviation of Salt Stress in Seedlings of Black Glutinous Rice by Seed Priming with Spermidine and Gibberellic Acid. Not. Bot. Horti Agrobot. Cluj-Napoca 2014, 42, 405–413. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, G.; Yang, W.; Wang, A.; Hu, Z.; Lin, C.; Chen, X. Drought-Inhibited Ribulose-1,5-Bisphosphate Carboxylase Activity Is Mediated through Increased Release of Ethylene and Changes in the Ratio of Polyamines in Pakchoi. J. Plant Physiol. 2014, 171, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- Gémes, K.; Mellidou, Ι.; Karamanoli, K.; Beris, D.; Park, K.Y.; Matsi, T.; Haralampidis, K.; Constantinidou, H.-I.; Roubelakis-Angelakis, K.A. Deregulation of Apoplastic Polyamine Oxidase Affects Development and Salt Response of Tobacco Plants. J. Plant Physiol. 2017, 211, 1–12. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, C.; Sun, C.; Wang, J.; Ye, Y.; Zhou, W.; Lu, L.; Lin, X. Inhibition of Ethylene Production by Putrescine Alleviates Aluminium-Induced Root Inhibition in Wheat Plants. Sci. Rep. 2016, 6, 18888. [Google Scholar] [CrossRef]
- Shah, A.A.; Ahmed, S.; Ali, A.; Yasin, N.A. 2-Hydroxymelatonin Mitigates Cadmium Stress in Cucumis sativus Seedlings: Modulation of Antioxidant Enzymes and Polyamines. Chemosphere 2020, 243, 125308. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Ashraf, M. Bioregulators: Unlocking Their Potential Role in Regulation of the Plant Oxidative Defense System. Plant Mol. Biol. 2021, 105, 11–41. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Polyamines and Abiotic Stress Tolerance in Plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef]
- Lenis, Y.Y.; Elmetwally, M.A.; Maldonado-Estrada, J.G.; Bazer, F.W. Physiological Importance of Polyamines. Zygote 2017, 25, 244–255. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Takahashi, Y.; Berberich, T.; Imai, A.; Takahashi, T.; Michael, A.J.; Kusano, T. A Protective Role for the Polyamine Spermine against Drought Stress in Arabidopsis. Biochem. Biophys. Res. Commun. 2007, 352, 486–490. [Google Scholar] [CrossRef]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine Is Involved in Arabidopsis Freezing Tolerance and Cold Acclimation by Regulating Abscisic Acid Levels in Response to Low Temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Shawky, B.T.; Ibrahim, A.S. Alleviation of Drought-Induced Oxidative Stress in Maize (Zea mays L.) Plants by Dual Application of 24-Epibrassinolide and Spermine. Environ. Exp. Bot. 2015, 113, 47–58. [Google Scholar] [CrossRef]
- Liu, J.; Yang, R.; Jian, N.; Wei, L.; Ye, L.; Wang, R.; Gao, H.; Zheng, Q. Putrescine Metabolism Modulates the Biphasic Effects of Brassinosteroids on Canola and Arabidopsis Salt Tolerance. Plant Cell Environ. 2020, 43, 1348–1359. [Google Scholar] [CrossRef]
- Van den Broeck, D.; Van Der Straeten, D.; Van Montagu, M.; Caplan, A. A Group of Chromosomal Proteins Is Specifically Released by Spermine and Loses DNA-Binding Activity upon Phosphorylation. Plant Physiol. 1994, 106, 559–566. [Google Scholar] [CrossRef]
- Childs, A.C.; Mehta, D.J.; Gerner, E.W. Polyamine-Dependent Gene Expression. Cell. Mol. Life Sci. 2003, 60, 1394–1406. [Google Scholar] [CrossRef]
- Du, H.; Chen, B.; Li, Q.; Liu, H.; Kurtenbach, R. Conjugated Polyamines in Root Plasma Membrane Enhanced the Tolerance of Plum Seedling to Osmotic Stress by Stabilizing Membrane Structure and Therefore Elevating H+-ATPase Activity. Front. Plant Sci. 2022, 12, 812360. [Google Scholar] [CrossRef]
- Kasukabe, Y.; He, L.; Nada, K.; Misawa, S.; Ihara, I.; Tachibana, S. Overexpression of Spermidine Synthase Enhances Tolerance to Multiple Environmental Stresses and Up-Regulates the Expression of Various Stress-Regulated Genes in Transgenic Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712–722. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Zhang, X.-Q.; Ma, X.; Huang, L.-K.; Yan, Y.-H. Exogenous Spermidine Improves Seed Germination of White Clover under Water Stress via Involvement in Starch Metabolism, Antioxidant Defenses and Relevant Gene Expression. Molecules 2014, 19, 18003–18024. [Google Scholar] [CrossRef]
- Sayed, A.I.; El-Hamahmy, M.A.M.; Rafudeen, M.S.; Ebrahim, M.K.H. Exogenous Spermidine Enhances Expression of Calvin Cycle Genes And Photosynthetic Efficiency in Sweet Sorghum Seedlings under Salt Stress. Biol. Plant. 2019, 63, 511–518. [Google Scholar] [CrossRef]
- Marco, F.; Busó, E.; Lafuente, T.; Carrasco, P. Spermine Confers Stress Resilience by Modulating Abscisic Acid Biosynthesis and Stress Responses in Arabidopsis Plants. Front. Plant Sci. 2019, 10, 972. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M.M.; Mahmud, J.-A.; Suzuki, T.; Fujita, M. Polyamines Confer Salt Tolerance in Mung Bean (Vigna radiata L.) by Reducing Sodium Uptake, Improving Nutrient Homeostasis, Antioxidant Defense, and Methylglyoxal Detoxification Systems. Front. Plant Sci. 2016, 7, 1104. [Google Scholar] [CrossRef]
- Bruggemann, L.I.; Pottosin, I.I.; Schonknecht, G. Cytoplasmic Polyamines Block the Fast-Activating Vacuolar Cation Channel. Plant J. 1998, 16, 101–105. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A.; Pottosin, I. Polyamines Prevent NaCl-Induced K+ Efflux from Pea Mesophyll by Blocking Non-Selective Cation Channels. FEBS Lett. 2007, 581, 1993–1999. [Google Scholar] [CrossRef] [PubMed]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium Signaling and Salt Tolerance Are Diversely Entwined in Plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef] [PubMed]
- Zepeda-Jazo, I.; Velarde-Buendía, A.M.; Enríquez-Figueroa, R.; Bose, J.; Shabala, S.; Muñiz-Murguía, J.; Pottosin, I.I. Polyamines Interact with Hydroxyl Radicals in Activating Ca2+ and K+ Transport across the Root Epidermal Plasma Membranes. Plant Physiol. 2011, 157, 2167–2180. [Google Scholar] [CrossRef]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Fuglsang, A.T.; Shabala, S. Polyamines Cause Plasma Membrane Depolarization, Activate Ca2+-, and Modulate H+-ATPase Pump Activity in Pea Roots. J. Exp. Bot. 2014, 65, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.P.; Kanwar, M.; Bhardwaj, R.; Yu, J.-Q.; Tran, L.-S.P. Chromium Stress Mitigation by Polyamine-Brassinosteroid Application Involves Phytohormonal and Physiological Strategies in Raphanus sativus L. PLoS ONE 2012, 7, e33210. [Google Scholar] [CrossRef]
- He, X.; Hao, J.; Fan, S.; Liu, C.; Han, Y. Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress. Plants 2022, 11, 1385. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Hassan, M.J.; Peng, Y.; Feng, G.; Huang, L.; Liu, L.; Liu, W.; Han, L.; Li, Z. Polyamines Metabolism Interacts with γ-Aminobutyric Acid, Proline and Nitrogen Metabolisms to Affect Drought Tolerance of Creeping Bentgrass. Int. J. Mol. Sci. 2022, 23, 2779. [Google Scholar] [CrossRef]
- Li, C.; Wang, G. Interactions between Reactive Oxygen Species, Ethylene and Polyamines in Leaves of Glycyrrhiza inflata Seedlings under Root Osmotic Stress. Plant Growth Regul. 2004, 42, 55–60. [Google Scholar] [CrossRef]
- Lee, T.-M.; Lur, H.-S.; Lin, Y.-H.; Chu, C. Physiological and Biochemical Changes Related to Methyl Jasmonate-Induced Chilling Tolerance of Rice (Oryza sativa L.) Seedlings. Plant Cell Environ. 1996, 19, 65–74. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, X.; Liu, W.; Kong, L.; Si, X.; Guo, S.; Sun, J. Gibberellin Mediates Spermidine-Induced Salt Tolerance and the Expression of GT-3b in Cucumber. Plant Physiol. Biochem. 2020, 152, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines Are Important in Abiotic Stress Signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Diao, Q.; Song, Y.; Shi, D.; Qi, H. Interaction of Polyamines, Abscisic Acid, Nitric Oxide, and Hydrogen Peroxide under Chilling Stress in Tomato (Lycopersicon esculentum Mill.) Seedlings. Front. Plant Sci. 2017, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Paschalidis, K.A.; Delis, I.D.; Andriopoulou, A.H.; Lagiotis, G.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Spermidine Exodus and Oxidation in the Apoplast Induced by Abiotic Stress Is Responsible for H2O2 Signatures That Direct Tolerance Responses in Tobacco. Plant Cell 2008, 20, 1708–1724. [Google Scholar] [CrossRef]
- Moschou, P.N.; Delis, I.D.; Paschalidis, K.A.; Roubelakis-Angelakis, K.A. Transgenic Tobacco Plants Overexpressing Polyamine Oxidase Are Not Able to Cope with Oxidative Burst Generated by Abiotic Factors. Physiol. Plant. 2008, 133, 140–156. [Google Scholar] [CrossRef]
- Rea, G.; de Pinto, M.C.; Tavazza, R.; Biondi, S.; Gobbi, V.; Ferrante, P.; De Gara, L.; Federico, R.; Angelini, R.; Tavladoraki, P. Ectopic Expression of Maize Polyamine Oxidase and Pea Copper Amine Oxidase in the Cell Wall of Tobacco Plants. Plant Physiol. 2004, 134, 1414–1426. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Li, J.; Li, C.; Xie, T.; Qu, D.; Meng, Y.; Li, C.; Wei, S. Exogenously Applied Spermidine Alleviates Photosynthetic Inhibition under Drought Stress in Maize (Zea mays L.) Seedlings Associated with Changes in Endogenous Polyamines and Phytohormones. Plant Physiol. Biochem. 2018, 129, 35–55. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, Y.-G. Plant Hormones in Salt Stress Tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between Plant Hormones and Heavy Metals Responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Sesin, V.; Kisiala, A.; Emery, R.J.N. Phytohormonal Roles in Plant Responses to Heavy Metal Stress: Implications for Using Macrophytes in Phytoremediation of Aquatic Ecosystems. Environ. Toxicol. Chem. 2021, 40, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Huang, B.; Jia, D.; Mann, T.; Jiang, X.; Qiu, Y.; Niitsu, M.; Berberich, T.; Kusano, T.; Liu, T. Identification of Seven Polyamine Oxidase Genes in Tomato (Solanum lycopersicum L.) and Their Expression Profiles under Physiological and Various Stress Conditions. J. Plant Physiol. 2018, 228, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yariuchi, Y.; Okamoto, T.; Noutoshi, Y.; Takahashi, T. Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings. Cells 2021, 10, 3283. [Google Scholar] [CrossRef]
- Takács, Z.; Czékus, Z.; Tari, I.; Poór, P. The Role of Ethylene Signalling in the Regulation of Salt Stress Response in Mature Tomato Fruits: Metabolism of Antioxidants and Polyamines. J. Plant Physiol. 2022, 277, 153793. [Google Scholar] [CrossRef]
- Alcázar, R.; García-Martínez, J.L.; Cuevas, J.C.; Tiburcio, A.F.; Altabella, T. Overexpression of ADC2 in Arabidopsis Induces Dwarfism and Late-Flowering through GA Deficiency: Overexpression of ADC2 in Arabidopsis. Plant J. 2005, 43, 425–436. [Google Scholar] [CrossRef]
- Ziosi, V.; Bregoli, A.M.; Bonghi, C.; Fossati, T.; Biondi, S.; Costa, G.; Torrigiani, P. Transcription of Ethylene Perception and Biosynthesis Genes Is Altered by Putrescine, Spermidine and Aminoethoxyvinylglycine (AVG) during Ripening in Peach Fruit (Prunus persica). New Phytol. 2006, 172, 229–238. [Google Scholar] [CrossRef]
- Tong, W.; Yoshimoto, K.; Kakehi, J.-I.; Motose, H.; Niitsu, M.; Takahashi, T. Thermospermine Modulates Expression of Auxin-Related Genes in Arabidopsis. Front. Plant Sci. 2014, 5, 94. [Google Scholar] [CrossRef]
- Aldesuquy, H.; Baka, Z.; Mickky, B. Kinetin and Spermine Mediated Induction of Salt Tolerance in Wheat Plants: Leaf Area, Photosynthesis and Chloroplast Ultrastructure of Flag Leaf at Ear Emergence. Egypt. J. Basic Appl. Sci. 2014, 1, 77–87. [Google Scholar] [CrossRef]
- Kapoor, R.T.; Hasanuzzaman, M. Exogenous Kinetin and Putrescine Synergistically Mitigate Salt Stress in Luffa acutangula by Modulating Physiology and Antioxidant Defense. Physiol. Mol. Biol. Plants 2020, 26, 2125–2137. [Google Scholar] [CrossRef]
- Xue, B.; Zhang, A.; Jiang, M. Involvement of Polyamine Oxidase in Abscisic Acid-Induced Cytosolic Antioxidant Defense in Leaves of Maize. J. Integr. Plant Biol. 2009, 51, 225–234. [Google Scholar] [CrossRef] [PubMed]
- He, M.-W.; Wang, Y.; Wu, J.-Q.; Shu, S.; Sun, J.; Guo, S.-R. Isolation and Characterization of S-Adenosylmethionine Synthase Gene from Cucumber and Responsive to Abiotic Stress. Plant Physiol. Biochem. 2019, 141, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Sagor, G.H.M.; Inoue, M.; Kusano, T.; Berberich, T. Expression Profile of Seven Polyamine Oxidase Genes in Rice (Oryza sativa) in Response to Abiotic Stresses, Phytohormones and Polyamines. Physiol. Mol. Biol. Plants 2021, 27, 1353–1359. [Google Scholar] [CrossRef]
- Tajti, J.; Hamow, K.Á.; Majláth, I.; Gierczik, K.; Németh, E.; Janda, T.; Pál, M. Polyamine-Induced Hormonal Changes in Eds5 and Sid2 Mutant Arabidopsis Plants. Int. J. Mol. Sci. 2019, 20, 5746. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Liu, J.; Liu, R.; Wu, H.; Jiang, C.; Wang, C.; Guan, Y. Temporal and Spatial Distributions of Sodium and Polyamines Regulated by Brassinosteroids in Enhancing Tomato Salt Resistance. Plant Soil 2016, 400, 147–164. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E.; Rorat, T.; Legocka, J. Polyamine Metabolism and S-Adenosylmethionine Decarboxylase Gene Expression during the Cytokinin-Stimulated Greening Process. Acta Physiol. Plant. 2007, 29, 495–502. [Google Scholar] [CrossRef]
- Cui, X.; Ge, C.; Wang, R.; Wang, H.; Chen, W.; Fu, Z.; Jiang, X.; Li, J.; Wang, Y. The BUD2 Mutation Affects Plant Architecture through Altering Cytokinin and Auxin Responses in Arabidopsis. Cell Res. 2010, 20, 576–586. [Google Scholar] [CrossRef]
- Parra-Lobato, M.C.; Gomez-Jimenez, M.C. Polyamine-Induced Modulation of Genes Involved in Ethylene Biosynthesis and Signalling Pathways and Nitric Oxide Production during Olive Mature Fruit Abscission. J. Exp. Bot. 2011, 62, 4447–4465. [Google Scholar] [CrossRef]
- Agudelo-Romero, P.; Ali, K.; Choi, Y.H.; Sousa, L.; Verpoorte, R.; Tiburcio, A.F.; Fortes, A.M. Perturbation of Polyamine Catabolism Affects Grape Ripening of Vitis vinifera Cv. Trincadeira. Plant Physiol. Biochem. 2014, 74, 141–155. [Google Scholar] [CrossRef]
- Dai, Y.-R.; Kaur-Sawhney, R.; Galston, A.W. Promotion by Gibberellic Acid of Polyamine Biosynthesis in Internodes of Light-Grown Dwarf Peas. Plant Physiol. 1982, 69, 103–106. [Google Scholar] [CrossRef]
- Shiozaki, S.; Ogata, T.; Horiuchi, S.; Zhuo, X. Involvement of Polyamines in Gibberellin-Induced Development of Seedless Grape Berries. Plant Growth Regul. 1998, 25, 187–193. [Google Scholar] [CrossRef]
- Kolotilin, I.; Koltai, H.; Bar-Or, C.; Chen, L.; Nahon, S.; Shlomo, H.; Levin, I.; Reuveni, M. Expressing Yeast SAMdc Gene Confers Broad Changes in Gene Expression and Alters Fatty Acid Composition in Tomato Fruit. Physiol. Plant. 2011, 142, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Zarza, X.; Atanasov, K.E.; Marco, F.; Arbona, V.; Carrasco, P.; Kopka, J.; Fotopoulos, V.; Munnik, T.; Gómez-Cadenas, A.; Tiburcio, A.F.; et al. Polyamine Oxidase 5 Loss-of-function Mutations in Arabidopsis thaliana Trigger Metabolic and Transcriptional Reprogramming and Promote Salt Stress Tolerance. Plant Cell Environ. 2017, 40, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.R.; Gárriz, A.; Marina, M.; Pieckenstain, F.L. Modulation of Polyamine Metabolism in Arabidopsis thaliana by Salicylic Acid. Physiol. Plant. 2021, 173, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Quinet, M.; Ndayiragije, A.; Lefevre, I.; Lambillotte, B.; Dupont-Gillain, C.C.; Lutts, S. Putrescine Differently Influences the Effect of Salt Stress on Polyamine Metabolism and Ethylene Synthesis in Rice Cultivars Differing in Salt Resistance. J. Exp. Bot. 2010, 61, 2719–2733. [Google Scholar] [CrossRef]
- Alcázar, R.; Bitrián, M.; Bartels, D.; Koncz, C.; Altabella, T.; Tiburcio, A.F. Polyamine Metabolic Canalization in Response to Drought Stress in Arabidopsis and the Resurrection Plant Craterostigma Plantagineum. Plant Signal. Behav. 2011, 6, 243–250. [Google Scholar] [CrossRef]
- Hou, Z.; Liu, G.; Hou, L.; Wang, L.; Liu, X. Regulatory Function of Polyamine Oxidase-Generated Hydrogen Peroxide in Ethylene-Induced Stomatal Closure in Arabidopsis thaliana. J. Integr. Agric. 2013, 12, 251–262. [Google Scholar] [CrossRef]
- Freitas, V.S.; de Souza Miranda, R.; Costa, J.H.; de Oliveira, D.F.; de Oliveira Paula, S.; de Castro Miguel, E.; Freire, R.S.; Prisco, J.T.; Gomes-Filho, E. Ethylene Triggers Salt Tolerance in Maize Genotypes by Modulating Polyamine Catabolism Enzymes Associated with H2O2 Production. Environ. Exp. Bot. 2018, 145, 75–86. [Google Scholar] [CrossRef]
- Moschou, P.N.; Sanmartin, M.; Andriopoulou, A.H.; Rojo, E.; Sanchez-Serrano, J.J.; Roubelakis-Angelakis, K.A. Bridging the Gap between Plant and Mammalian Polyamine Catabolism: A Novel Peroxisomal Polyamine Oxidase Responsible for a Full Back-Conversion Pathway in Arabidopsis. Plant Physiol. 2008, 147, 1845–1857. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Islam, M.S.; Hossain, A.; Iqbal, M.A.; Mubeen, M.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as Growth Regulators During Abiotic Stress Tolerance in Plants. Front. Agron. 2022, 4, 765068. [Google Scholar] [CrossRef]
- An, Z.; Jing, W.; Liu, Y.; Zhang, W. Hydrogen Peroxide Generated by Copper Amine Oxidase Is Involved in Abscisic Acid-Induced Stomatal Closure in Vicia faba. J. Exp. Bot. 2008, 59, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Fincato, P.; Moschou, P.N.; Ahou, A.; Angelini, R.; Roubelakis-Angelakis, K.A.; Federico, R.; Tavladoraki, P. The Members of Arabidopsis thaliana PAO Gene Family Exhibit Distinct Tissue- and Organ-Specific Expression Pattern during Seedling Growth and Flower Development. Amino Acids 2012, 42, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Polyamines Increase Nitric Oxide and Reactive Oxygen Species in Guard Cells of Arabidopsis thaliana during Stomatal Closure. Protoplasma 2018, 255, 153–162. [Google Scholar] [CrossRef]
- Alcázar, R.; Cuevas, J.C.; Patron, M.; Altabella, T.; Tiburcio, A.F. Abscisic Acid Modulates Polyamine Metabolism under Water Stress in Arabidopsis thaliana. Physiol. Plant. 2006, 128, 448–455. [Google Scholar] [CrossRef]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of Polyamines in Plant Response to Abiotic Stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A.; Sengupta, D.N. Identification of Trans-Acting Factors Regulating SamDC Expression in Oryza sativa. Biochem. Biophys. Res. Commun. 2014, 445, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-D.; Zhang, M.; Gao, D.-J.; Zhou, K.; Tang, S.-J.; Zhou, B.; Lv, Y.-M. Rice OsHSFA3 Gene Improves Drought Tolerance by Modulating Polyamine Biosynthesis Depending on Abscisic Acid and ROS Levels. Int. J. Mol. Sci. 2020, 21, 1857. [Google Scholar] [CrossRef] [PubMed]
- Espasandin, F.D.; Maiale, S.J.; Calzadilla, P.; Ruiz, O.A.; Sansberro, P.A. Transcriptional Regulation of 9-Cis-Epoxycarotenoid Dioxygenase (NCED) Gene by Putrescine Accumulation Positively Modulates ABA Synthesis and Drought Tolerance in Lotus tenuis Plants. Plant Physiol. Biochem. 2014, 76, 29–35. [Google Scholar] [CrossRef]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of Polyamines, Abscisic Acid and Proline under Osmotic Stress in the Leaves of Wheat Plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef]
- Marcińska, I.; Dziurka, K.; Waligórski, P.; Janowiak, F.; Skrzypek, E.; Warchoł, M.; Juzoń, K.; Kapłoniak, K.; Czyczyło-Mysza, I.M. Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress. Life 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, H.; Lv, X.; Liu, D.; Wen, X.; Liao, Y. Effect of polyamines on the grain filling of wheat under drought stress. Plant Physiol. Biochem. 2016, 100, 113–129. [Google Scholar] [CrossRef]
- Peremarti, A.; Bassie, L.; Yuan, D.; Pelacho, A.; Christou, P.; Capell, T. Transcriptional Regulation of the Rice Arginine Decarboxylase (Adc1) and S-Adenosylmethionine Decarboxylase (Samdc) Genes by Methyl Jasmonate. Plant Physiol. Biochem. 2010, 48, 553–559. [Google Scholar] [CrossRef]
- Krishnan, S.; Merewitz, E.B. Polyamine Application Effects on Gibberellic Acid Content in Creeping Bentgrass during Drought Stress. J. Am. Soc. Hortic. Sci. 2017, 142, 135–142. [Google Scholar] [CrossRef]
- Serna, M.; Coll, Y.; Zapata, P.J.; Botella, M.Á.; Pretel, M.T.; Amorós, A. A Brassinosteroid Analogue Prevented the Effect of Salt Stress on Ethylene Synthesis and Polyamines in Lettuce Plants. Sci. Hortic. 2015, 185, 105–112. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Lee, I.-J. Regulation of Salicylic Acid, Jasmonic Acid and Fatty Acids in Cucumber (Cucumis sativus L.) by Spermidine Promotes Plant Growth against Salt Stress. Acta Physiol. Plant. 2013, 35, 3315–3322. [Google Scholar] [CrossRef]
- Alsokari, S.S. Synergistic Effect of Kinetin and Spermine on Some Physiological Aspects of Seawater Stressed Vigna sinensis Plants. Saudi J. Biol. Sci. 2011, 18, 37–44. [Google Scholar] [CrossRef]
- Kaur, H.; Bhardwaj, R.; Thukral, A.K. Modulation of Jasmonic Acid and Polyamines by 24- Epibrassinolide in Brassica juncea L. under Copper Stress. IOSR J. Environ. Sci. Toxicol. Food Technol. 2016, 10, 1–12. [Google Scholar] [CrossRef]
- Kohli, S.K.; Bali, S.; Tejpal, R.; Bhalla, V.; Verma, V.; Bhardwaj, R.; Alqarawi, A.A.; Abd Allah, E.F.; Ahmad, P. In-Situ Localization and Biochemical Analysis of Bio-Molecules Reveals Pb-Stress Amelioration in Brassica juncea L. by Co-Application of 24-Epibrassinolide and Salicylic Acid. Sci. Rep. 2019, 9, 3524. [Google Scholar] [CrossRef]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine as a Signal to Modulate the Indispensable ABA Increase under Cold Stress. Plant Signal. Behav. 2009, 4, 219–220. [Google Scholar] [CrossRef]
- Wang, C.Y.; Buta, J.G. Methyl Jasmonate Reduces Chilling Injury in Cucurbita pepo through Its Regulation of Abscisic Acid and Polyamine Levels. Environ. Exp. Bot. 1994, 34, 427–432. [Google Scholar] [CrossRef]
- Zeng, Y.; Zahng, Y.; Xiang, J.; Wu, H.; Chen, H.; Zhang, Y.; Zhu, D. Effects of Chilling Tolerance Induced by Spermidine Pretreatment on Antioxidative Activity, Endogenous Hormones and Ultrastructure of Indica-Japonica Hybrid Rice Seedlings. J. Integr. Agric. 2016, 15, 295–308. [Google Scholar] [CrossRef]
- Gomez-Cadenas, A.; Vives, V.; Zandalinas, S.; Manzi, M.; Sanchez-Perez, A.; Perez-Clemente, R.; Arbona, V. Abscisic Acid: A Versatile Phytohormone in Plant Signaling and Beyond. Curr. Protein Pept. Sci. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Hama, J.R.; Hooshmand, K.; Laursen, B.B.; Vestergård, M.; Fomsgaard, I.S. Clover Root Uptake of Cereal Benzoxazinoids (BXs) Caused Accumulation of BXs and BX Transformation Products Concurrently with Substantial Increments in Clover Flavonoids and Abscisic Acid. J. Agric. Food Chem. 2022, 70, 14633–14640. [Google Scholar] [CrossRef] [PubMed]
- Murcia, G.; Fontana, A.; Pontin, M.; Baraldi, R.; Bertazza, G.; Piccoli, P.N. ABA and GA3 Regulate the Synthesis of Primary and Secondary Metabolites Related to Alleviation from Biotic and Abiotic Stresses in Grapevine. Phytochemistry 2017, 135, 34–52. [Google Scholar] [CrossRef]
- Song, Y.; Miao, Y.; Song, C. Behind the Scenes: The Roles of Reactive Oxygen Species in Guard Cells. New Phytol. 2014, 201, 1121–1140. [Google Scholar] [CrossRef]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Rodrigues-Pousada, R.A.; Cona, A. Developmental, Hormone- and Stress-Modulated Expression Profiles of Four Members of the Arabidopsis Copper-Amine Oxidase Gene Family. Plant Physiol. Biochem. 2020, 147, 141–160. [Google Scholar] [CrossRef]
- Marin, E.; Nussaume, L.; Quesada, A.; Gonneau, M.; Sotta, B.; Hugueney, P.; Frey, A.; Marion-Poll, A. Molecular Identification of Zeaxanthin Epoxidase of Nicotiana plumbaginifolia, a Gene Involved in Abscisic Acid Biosynthesis and Corresponding to the ABA Locus of Arabidopsis thaliana. EMBO J. 1996, 15, 2331–2342. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Tan, B.C.; Gage, D.A.; Zeevaart, J.A.D.; McCarty, D.R. Specific Oxidative Cleavage of Carotenoids by VP14 of Maize. Science 1997, 276, 1872–1874. [Google Scholar] [CrossRef]
- Seo, M.; Koiwai, H.; Akaba, S.; Komano, T.; Oritani, T.; Kamiya, Y.; Koshiba, T. Abscisic Aldehyde Oxidase in Leaves of Arabidopsis thaliana. Plant J. 2000, 23, 481–488. [Google Scholar] [CrossRef]
- Konstantinos, P.A.; Imene, T.; Panagiotis, M.N.; Roubelakis-Angelakis, K.A. ABA-Dependent Amine Oxidases-Derived H2O2 Affects Stomata Conductance. Plant Signal. Behav. 2010, 5, 1153–1156. [Google Scholar] [CrossRef]
- Toumi, I.; Moschou, P.N.; Paschalidis, K.A.; Bouamama, B.; Ben Salem-fnayou, A.; Ghorbel, A.W.; Mliki, A.; Roubelakis-Angelakis, K.A. Abscisic Acid Signals Reorientation of Polyamine Metabolism to Orchestrate Stress Responses via the Polyamine Exodus Pathway in Grapevine. J. Plant Physiol. 2010, 167, 519–525. [Google Scholar] [CrossRef]
- Skowron, E.; Trojak, M. Effect of Exogenously-Applied Abscisic Acid, Putrescine and Hydrogen Peroxide on Drought Tolerance of Barley. Biologia 2021, 76, 453–468. [Google Scholar] [CrossRef]
- Hasan, M.M.; Skalicky, M.; Jahan, M.S.; Hossain, M.N.; Anwar, Z.; Nie, Z.-F.; Alabdallah, N.M.; Brestic, M.; Hejnak, V.; Fang, X.-W. Spermine: Its Emerging Role in Regulating Drought Stress Responses in Plants. Cells 2021, 10, 261. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Slathia, S.; Choudhary, S.P.; Sharma, Y.P.; Langer, A. Role of 24-Epibrassinolide, Putrescine and Spermine in Salinity Stressed Adiantum Capillus-Veneris Leaves. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 183–192. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. Jasmonates: Mechanisms and Functions in Abiotic Stress Tolerance of Plants. Biocatal. Agric. Biotechnol. 2019, 20, 101210. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, M.I.R.; Anjum, N.A.; Masood, A.; Hussain, S.J.; Khan, N.A. Jasmonates in Plants under Abiotic Stresses: Crosstalk with Other Phytohormones Matters. Environ. Exp. Bot. 2018, 145, 104–120. [Google Scholar] [CrossRef]
- Noir, S.; Bömer, M.; Takahashi, N.; Ishida, T.; Tsui, T.-L.; Balbi, V.; Shanahan, H.; Sugimoto, K.; Devoto, A. Jasmonate Controls Leaf Growth by Repressing Cell Proliferation and the Onset of Endoreduplication While Maintaining a Potential Stand-By Mode. Plant Physiol. 2013, 161, 1930–1951. [Google Scholar] [CrossRef]
- Attaran, E.; Major, I.T.; Cruz, J.A.; Rosa, B.A.; Koo, A.J.K.; Chen, J.; Kramer, D.M.; He, S.Y.; Howe, G.A. Temporal Dynamics of Growth and Photosynthesis Suppression in Response to Jasmonate Signaling. Plant Physiol. 2014, 165, 1302–1314. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Sidhu, G.P.S.; Kumar, R.; Kohli, S.K.; Yadav, P.; Kapoor, D.; Bali, A.S.; Shahzad, B.; Khanna, K.; et al. Abiotic Stress Management in Plants: Role of Ethylene. In Molecular Plant Abiotic Stress; Roychoudhury, A., Tripathi, D., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 185–208. ISBN 978-1-119-46366-5. [Google Scholar]
- Grzesiak, M.; Filek, M.; Barbasz, A.; Kreczmer, B.; Hartikainen, H. Relationships between Polyamines, Ethylene, Osmoprotectants and Antioxidant Enzymes Activities in Wheat Seedlings after Short-Term PEG- and NaCl-Induced Stresses. Plant Growth Regul. 2013, 69, 177–189. [Google Scholar] [CrossRef]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Muneeb-u-Rehman; Siddiqi, T.O.; Ahmad, P. Salt Stress: Causes Types and Responses of Plants. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 1–24. ISBN 978-1-4614-4746-7. [Google Scholar]
- Ahmad, P.; Abass Ahanger, M.; Nasser Alyemeni, M.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of Sodium Chloride Toxicity in Solanum lycopersicum L. by Supplementation of Jasmonic Acid and Nitric Oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef]
- Zapata, P.J.; Serrano, M.; García-Legaz, M.F.; Pretel, M.T.; Botella, M.A. Short Term Effect of Salt Shock on Ethylene and Polyamines Depends on Plant Salt Sensitivity. Front. Plant Sci. 2017, 8, 855. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.F.; Hoffman, N.E. Ethylene Biosynthesis and Its Regulation in Higher Plants. Annu. Rev. Plant. Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Zhong, S.; Lin, Z.; Grierson, D. Tomato Ethylene Receptor–CTR Interactions: Visualization of NEVER-RIPE Interactions with Multiple CTRs at the Endoplasmic Reticulum. J. Exp. Bot. 2008, 59, 965–972. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Mir, B.A.; Yusuf, M.; Ahmad, A. 24-Epibrassinolide and/or Putrescine Trigger Physiological and Biochemical Responses for the Salt Stress Mitigation in Cucumis sativus L. Photosynthetica 2014, 52, 464–474. [Google Scholar] [CrossRef]
- Pál, M.; Ivanovska, B.; Oláh, T.; Tajti, J.; Hamow, K.Á.; Szalai, G.; Khalil, R.; Vanková, R.; Dobrev, P.; Misheva, S.P.; et al. Role of Polyamines in Plant Growth Regulation of Rht Wheat Mutants. Plant Physiol. Biochem. 2019, 137, 189–202. [Google Scholar] [CrossRef]
- Pavlů, J.; Novák, J.; Koukalová, V.; Luklová, M.; Brzobohatý, B.; Černý, M. Cytokinin at the Crossroads of Abiotic Stress Signalling Pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef]
- Hussain, A.; Nazir, F.; Fariduddin, Q. 24-Epibrassinolide and Spermidine Alleviate Mn Stress via the Modulation of Root Morphology, Stomatal Behavior, Photosynthetic Attributes and Antioxidant Defense in Brassica juncea. Physiol. Mol. Biol. Plants 2019, 25, 905–919. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Oral, H.V.; Bhardwaj, R.; Yu, J.-Q.; Tran, L.-S.P. Interaction of Brassinosteroids and Polyamines Enhances Copper Stress Tolerance in Raphanus sativus. J. Exp. Bot. 2012, 63, 5659–5675. [Google Scholar] [CrossRef]
- Li, M.; Duan, X.; Gao, G.; Liu, T.; Qi, H. CmABF1 and CmCBF4 Cooperatively Regulate Putrescine Synthesis to Improve Cold Tolerance of Melon Seedlings. Hortic. Res. 2022, 9, uhac002. [Google Scholar] [CrossRef]
- Lang, V.; Mantyla, E.; Welin, B.; Sundberg, B.; Palva, E.T. Alterations in Water Status, Endogenous Abscisic Acid Content, and Expression of Rab18 Gene during the Development of Freezing Tolerance in Arabidopsis thaliana. Plant Physiol. 1994, 104, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- González-Aguilar, G.A.; Fortiz, J.; Cruz, R.; Baez, R.; Wang, C.Y. Methyl Jasmonate Reduces Chilling Injury and Maintains Postharvest Quality of Mango Fruit. J. Agric. Food Chem. 2000, 48, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Honda, C.; Kondo, S. Effect of Low-Temperature Stress on Abscisic Acid, Jasmonates, and Polyamines in Apples. Plant Growth Regul. 2007, 52, 199–206. [Google Scholar] [CrossRef]
- Peng, J.; Li, Z.; Wen, X.; Li, W.; Shi, H.; Yang, L.; Zhu, H.; Guo, H. Salt-Induced Stabilization of EIN3/EIL1 Confers Salinity Tolerance by Deterring ROS Accumulation in Arabidopsis. PLoS Genet. 2014, 10, e1004664. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, E.; Martínez, J.-P.; Benahmed, H.; Lepoint, G.; Vanpee, B.; Quinet, M.; Lutts, S. Inhibition of Ethylene Synthesis Reduces Salt-Tolerance in Tomato Wild Relative Species Solanum Chilense. J. Plant Physiol. 2017, 210, 24–37. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Nawaz, F.; Shabbir, R.N.; Shahbaz, M.; Majeed, S.; Raheel, M.; Hassan, W.; Sohail, M.A. Cross Talk between Nitric Oxide and Phytohormones Regulate Plant Development during Abiotic Stresses. In Phytohormones—Signaling Mechanisms and Crosstalk in Plant Development and Stress Responses; El-Esawi, M., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-3411-4. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).