

Stability Evaluation for Heat Tolerance in Lettuce: Implications and Recommendations


Abstract
:1. Introduction
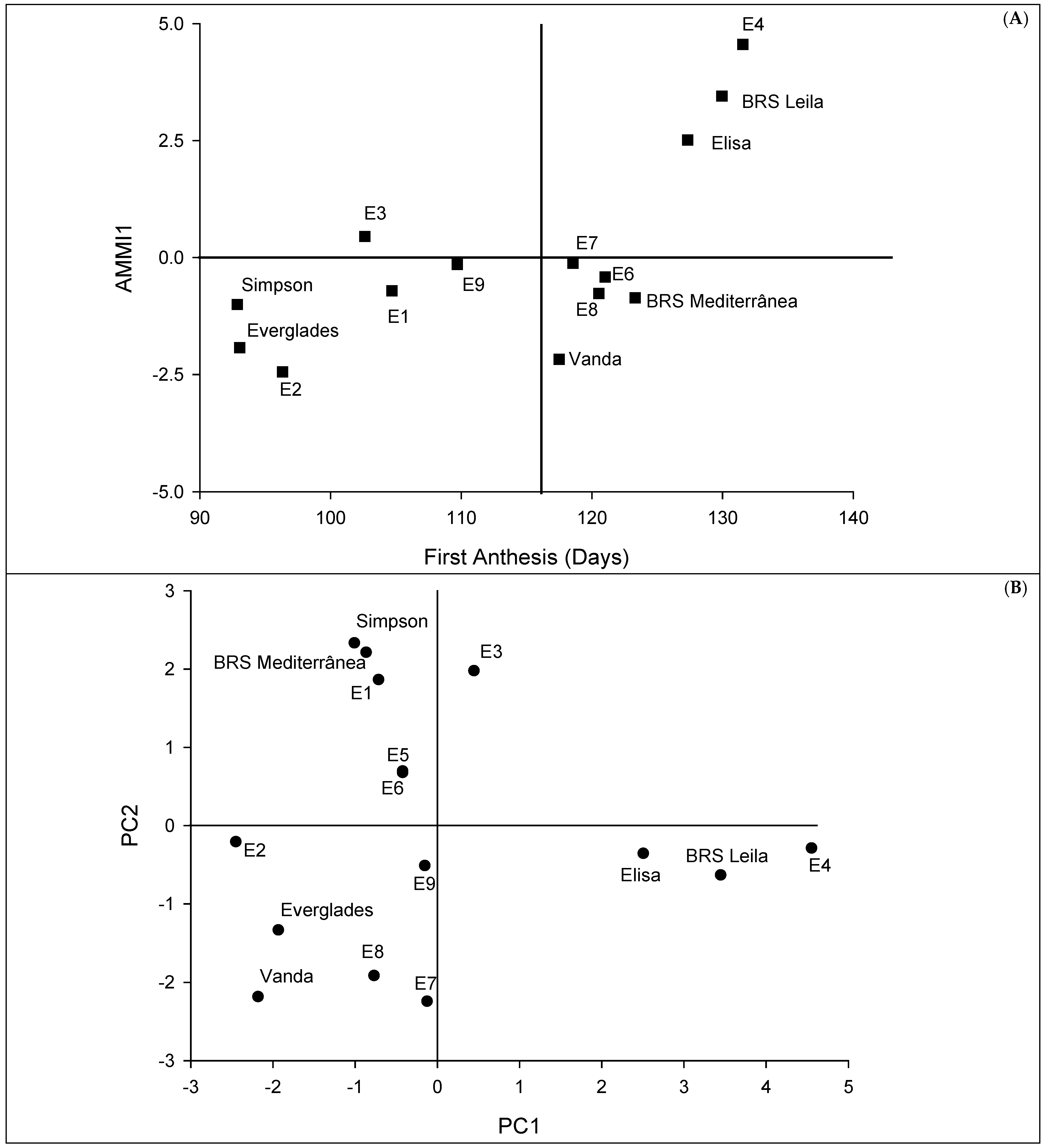
2. Results
3. Discussion
4. Materials and Methods
4.1. General Conditions
4.2. Experimental Design and Data Collection
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Lettuce Type | Genotypes | Origin | Pedigree | Breeding Programam |
|---|---|---|---|---|
| Leaf | BRS Leila | Brazil | CNPH 473/CNPH 12 | EMBRAPA/Agrocinco DUX Co. (Monte Mor, SP, Brazil) |
| BRS Mediterrânea | Brazil | CNPH 287/CNPH 49 | EMBRAPA/Agrocinco DUX Co. | |
| Simpson | USA | Unavailable | Henderson Seed Company (Melbourne, Australia) | |
| Vanda | Brazil | Unavailable | Sakata Seeds Co. (Yokohama, Japan) | |
| Butterhead | Elisa | Brazil | Unavailable | Sakata Seeds Co. |
| Crisphead | Everglades | USA | Great Lakes/Fulton/Gallega/Dark Green Boston/Troc | University of Florida (Gainesville, FL, USA) |
References
- Haytowitz, D.B.; Lemar, L.E.; Pehrsson, P.R. USDA’s Nutrient Databank System—A Tool for Handling Data from Diverse Sources. J. Food Compos. Anal. 2009, 22, 433–441. [Google Scholar] [CrossRef]
- Pitrat, M. Vegetable Crops in the Mediterranean Basin with an Overview of Virus Resistance. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2012; Volume 84, pp. 1–29. ISBN 978-0-12-394314-9. [Google Scholar]
- Wei, S.; Zhang, L.; Huo, G.; Ge, G.; Luo, L.; Yang, Q.; Yang, X.; Long, P. Comparative Transcriptomics and Metabolomics Analyses Provide Insights into Thermal Resistance in Lettuce (Lactuca sativa L.). Sci. Hortic. 2021, 289, 110423. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.; Wang, J. Effects of Light Intensity and Temperature on the Photosynthesis Characteristics and Yield of Lettuce. Horticulturae 2022, 8, 178. [Google Scholar] [CrossRef]
- Jenni, S.; Yan, W. Genotype by Environment Interactions of Heat Stress Disorder Resistance in Crisphead Lettuce. Plant Breed. 2009, 128, 374–380. [Google Scholar] [CrossRef]
- Lafta, A.; Sandoya, G.; Mou, B. Genetic Variation and Genotype by Environment Interaction for Heat Tolerance in Crisphead Lettuce. HortScience Horts 2021, 56, 126–135. [Google Scholar] [CrossRef]
- Collado-González, J.; Piñero, M.C.; Otalora, G.; López-Marín, J.; Del Amor, F.M. Assessing Optimal Nitrate/Ammonium-Ratios in Baby-Leaf Lettuce to Enhance the Heat Stress Tolerance under Elevated CO2 Conditions. PLoS ONE 2022, 17, e0278309. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Razzaq, A.; Mehmood, S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Legg, S. IPCC, 2021: Climate Change 2021-the Physical Science Basis. Interaction 2021, 49, 44–45. [Google Scholar]
- Chandra, K.; Chand, S.; Saini, R.P.; Sharma, R. (Eds.) Smart Breeding: Molecular Interventions and Advancements for Crop Improvement; Apple Academic Press: Palm Bay, FL, USA; CRC Press: Burlington, ON, Canada, 2023; ISBN 978-1-00-336186-2. [Google Scholar]
- Yue, H.; Gauch, H.G.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Bu, J.; Jiang, X. Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China. Agriculture 2022, 12, 602. [Google Scholar] [CrossRef]
- Diouf, I.; Derivot, L.; Koussevitzky, S.; Carretero, Y.; Bitton, F.; Moreau, L.; Causse, M. Genetic Basis of Phenotypic Plasticity and Genotype × Environment Interactions in a Multi-Parental Tomato Population. J. Exp. Bot. 2020, 71, 5365–5376. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G.; Ierna, A. Dissecting the Genotype × Environment Interaction for Potato Tuber Yield and Components. Agronomy 2022, 13, 101. [Google Scholar] [CrossRef]
- Yang, X.; Soliman, A.A.; Hu, C.; Yang, F.; Lv, M.; Yu, H.; Wang, Y.; Zheng, A.; Dai, Z.; Li, Q.; et al. Yield Adaptability and Stability in Field Pea Genotypes Using AMMI, GGE, and GYT Biplot Analyses. Agriculture 2023, 13, 1962. [Google Scholar] [CrossRef]
- Yahaya, M.A.; Shimelis, H.; Nebié, B.; Mashilo, J.; Pop, G. Response of African Sorghum Genotypes for Drought Tolerance under Variable Environments. Agronomy 2023, 13, 557. [Google Scholar] [CrossRef]
- Becker, H.C.; Leon, J.I. Stability Analysis in Plant Breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Khalili, M.; Poczai, P.; Olivoto, T. Stability Indices to Deciphering the Genotype-by-Environment Interaction (GEI) Effect: An Applicable Review for Use in Plant Breeding Programs. Plants 2022, 11, 414. [Google Scholar] [CrossRef] [PubMed]
- Wricke, G. Uber Eine Methode Zur Erfassung Der Okologischen Streubreite in Feldversucen. Z. Pflanzenzucht. 1962, 47, 92–96. [Google Scholar]
- Eberhart, S.A.; Russell, W.A. Stability Parameters for Comparing Varieties 1. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G.N. The Analysis of Adaptation in a Plant-Breeding Programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef]
- Cruz, C.D. Modelos Biometricos Aplicados Ao Melhoramento Geneticos; Editora UFV: Vancouver, BC, Canada, 2002; ISBN 978-85-7269-151-2. [Google Scholar]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical Analysis of a Yield Trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C.; Da Silva, J.A.G.; Marchioro, V.S.; De Souza, V.Q.; Jost, E. Mean Performance and Stability in Multi-Environment Trials I: Combining Features of AMMI and BLUP Techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- Annicchiarico, P. Cultivar Adaptation and Recommendation from Alfalfa Trials in Northern Italy. J. Genet. Breed. 1992, 46, 269. [Google Scholar]
- Huehn, M. Nonparametric Measures of Phenotypic Stability. Part 2: Applications. Euphytica 1990, 47, 195–201. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R. A superiority measure of cultivar performance for cultivar × location data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Carneiro, P.C.S. Novas Metodologias de Análise Da Adaptabilidade e Estabilidade de Comportamento; Universidade Federal de Viçosa: Viçosa, MG, Brazil, 1998. [Google Scholar]
- Scott, A.J.; Knott, M. A Cluster Analysis Method for Grouping Means in the Analysis of Variance. Biometrics 1974, 30, 507–512. [Google Scholar] [CrossRef]
- Liu, R.; Su, Z.; Zhou, H.; Huang, Q.; Fan, S.; Liu, C.; Han, Y. LsHSP70 Is Induced by High Temperature to Interact with Calmodulin, Leading to Higher Bolting Resistance in Lettuce. Sci. Rep. 2020, 10, 15155. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Han, Y.; Ning, K.; Ding, Y.; Zhao, W.; Yan, S.; Luo, C.; Jiang, X.; Ge, D.; Liu, R. Inflorescence Development and the Role of LsFT in Regulating Bolting in Lettuce (Lactuca sativa L.). Front. Plant Sci. 2018, 8, 2248. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, W.; Ge, D.; Han, Y.; Ning, K.; Luo, C.; Wang, S.; Liu, R.; Zhang, X.; Wang, Q. LCM-seq Reveals the Crucial Role of LsSOC1 in Heat-promoted Bolting of Lettuce (Lactuca sativa L.). Plant J. 2018, 95, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Abbass, K.; Qasim, M.Z.; Song, H.; Murshed, M.; Mahmood, H.; Younis, I. A Review of the Global Climate Change Impacts, Adaptation, and Sustainable Mitigation Measures. Environ. Sci. Pollut. Res. 2022, 29, 42539–42559. [Google Scholar] [CrossRef]
- Oroian, C.; Ugruțan, F.; Mureșan, I.C.; Oroian, I.; Odagiu, A.; Petrescu-Mag, I.V.; Burduhos, P. AMMI Analysis of Genotype × Environment Interaction on Sugar Beet (Beta vulgaris L.) Yield, Sugar Content and Production in Romania. Agronomy 2023, 13, 2549. [Google Scholar] [CrossRef]
- Aditya, J.P.; Bhartiya, P.; Bhartiya, A. Genetic Variability, Heritability and Character Association for Yield and Component Characters in Soybean (G. Max (L.) Merrill). J. Cent. Eur. Agric. 2011, 12, 27–34. [Google Scholar] [CrossRef]
- Silva, J.L.T.; Cavalcante, M.; Barros, J.A.S. Estabilidade e Adaptabilidade de Cultivares de Alface Em Diferentes Substratos. Comun. Sci. 2022, 14, e3886. [Google Scholar] [CrossRef]
- Gualberto, R.; de Oliveira, P.S.R.; Guimarães, A. de M. Adaptabilidade e Estabilidade Fenotípica de Cultivares de Alface Do Grupo Crespa Em Cultivo Hidropônico. Hortic. Bras. 2009, 27, 7–11. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Barati, A.; Koohkan, S.A.; Jabari, M.; Marzoghian, A.; Gholipoor, A.; Shahbazi-Homonloo, K.; Zali, H.; Poodineh, O.; Kheirgo, M. Dissection of Genotype-by-Environment Interaction and Yield Stability Analysis in Barley Using AMMI Model and Stability Statistics. Bull. Natl. Res. Cent. 2022, 46, 19. [Google Scholar] [CrossRef]
- Scapim, C.A.; Pacheco, C.A.P.; do Amaral Júnior, A.T.; Vieira, R.A.; Pinto, R.J.B.; Conrado, T.V. Correlations between the Stability and Adaptability Statistics of Popcorn Cultivars. Euphytica 2010, 174, 209–218. [Google Scholar] [CrossRef]
- Schmildt, E.R.; Cruz, C.D. Análise da adaptabilidade e estabilidade do milho pelos métodos de eberhart e russell e de annicchiarico/adaptability and stability of maize using eberhart/russell and annicchiarico methods. Ceres 2005, 52, 45–58. [Google Scholar]
- Guzman, V.L.; Nagata, R.T.; Datnoff, L.E.; Raid, R.N. “Florida 202’and’Everglades”: Two New Butterhead Lettuce Cultivars Adapted to Florida. HortScience 1992, 27, 852–853. [Google Scholar] [CrossRef]
- Cajamarca, S.M.N.; Lima, C.E.P.; Silva, J.D.; Guedes, I.M.R.; Braga, M.B.; Fontenelle, M.R.; Figueiredo, C.C.D. Curly Lettuce Development, Nutrient Absorption and Salinization Tolerance in Response to an Aerobic Biofertilizer Produced from Agro-Industrial Residues. Aust. J. Crop Sci. 2019, 13, 1659–1667. [Google Scholar] [CrossRef]
- Suinaga, F.A.; Resende, F.; Boiteux, L.S.; Pinheiro, J.B. Potencial Produtivo de Linhagens de Alface Crespa: II-Cultivo Orgânico; Embrapa Hortaliças: Brasilia, DF, Brazil, 2014. [Google Scholar]
- Suinaga, F.A.; Souza, N.O.S.; Boiteux, L.S.; Pinheiro, J.B. Potencial Produtivo de Linhagens de Alface Crespa: I–Vargem Bonita-Boletim Pesquisa e Desenvolvimento/Embrapa Hortaliças; Embrapa Hortaliças: Brasília, DF, Brazil, 2014. [Google Scholar]
- Shukla, G.K. Some Statistical Aspects of Partitioning Genotype-Environmental Components of Variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef]
- SAS. SAS System, Version 9.1. 3; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Olivoto, T.; Lúcio, A.D. Metan: An R Package for Multi-environment Trial Analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Cruz, C.D. Programa Genes-Ampliado e Integrado Aos Aplicativos R, Matlab e Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]

| Environments (E) | Year | Season | Place | Average Temperature (°C) |
|---|---|---|---|---|
| E1 | 2015 | Summer | Field | 24 |
| E2 | 2015 | Summer | Glass Greenhouse | 30 |
| E3 | 2015 | Summer | Plastic Greenhouse | 27 |
| E4 | 2016 | Fall | Field | 20 |
| E5 | 2016 | Fall | Glass Greenhouse | 27 |
| E6 | 2016 | Fall | Plastic Greenhouse | 25 |
| E7 | 2016 | Winter | Field | 24 |
| E8 | 2016 | Winter | Glass Greenhouse | 29 |
| E9 | 2016 | Winter | Plastic Greenhouse | 26 |
| Source of Variation | DF | SS | MS | Cumulative (%) |
|---|---|---|---|---|
| Replication/Environment | 27 | 2390.04 | 88.52 | |
| Environment | 8 | 24,369.28 | 3046.16 ** | |
| Genotype | 5 | 50,989.30 | 10,197.86 ** | |
| Genotype × Environment | 40 | 5706.40 | 142.66 ** | |
| PC1 | 12 | 3232.20 | 269.35 * | 56.60 |
| PC2 | 10 | 1213.10 | 121.31 * | 77.90 |
| PC3 | 8 | 999.28 | 124.91 * | 95.40 |
| PC4 | 6 | 205.80 | 34.30 | 99.00 |
| PC5 | 4 | 56.16 | 14.04 | 100.00 |
| Genotype × Environment (linear) | 5 | 1794.10 | 358.82 ** | |
| Deviation | 42 | 3912.30 | 93.15 ** | |
| Error | 135 | 1464.75 | 10.85 |
| Genotypes | Environments 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| E1 | E2 | E3 | E4 | E5 | E6 | E7 | E8 | E9 | |
| BRS Leila | 116 A | 106 A | 126 A | 163 A | 134 A | 131 A | 137 A | 137 A | 123 A |
| BRS Mediterrânea | 115 A | 106 A | 120 A | 135 B | 128 B | 133 A | 124 B | 127 C | 117 B |
| Elisa | 117 A | 102 A | 108 B | 157 A | 121 C | 138 A | 131 B | 130 B | 126 A |
| Everglades | 84 D | 81 B | 77 C | 102 D | 97 E | 100 C | 103 C | 101 D | 90 C |
| Simpson Black Seed | 92 C | 78 B | 83 C | 106 D | 89 F | 101 C | 91 D | 96 E | 89 C |
| Vanda | 105 B | 105 A | 103 B | 126 C | 117 D | 124 B | 126 B | 132 B | 114 B |
| Average | 105 | 96 | 103 | 132 | 114 | 121 | 119 | 121 | 110 |
| CV (%) | 1.70 | 2.70 | 6.02 | 2.74 | 1.52 | 2.27 | 2.54 | 1.25 | 2.91 |
| Environmental Index 2 | −9.30 | −17.67 | −11.38 | 17.54 | 7.00 | 7.20 | 4.54 | 6.54 | −4.29 |
| Genotypes | First Anthesis (Days) | Eberhardt and Russell | Lin and Binns Modified by Carneiro (Pi’s/10,000) | Wricke’s Ecovalence | Shukla’s Stability Variance | WAASB | Mean Rank | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | Pi | Pi+ | Pi− | ||||||||
| BRS Leila | 130 (1) | 1.30 ** | 46.51 ** (6) | 83.24 | 4.76 (1) | 3.21 (1) | 1.54 (1) | 285.46 (6) | 1739.09 (6) | 2.42 (6) | 3.5 |
| BRS Mediterrânea | 123 (3) | 0.80 * | 12.18 ** (2) | 86.21 | 7.11 (3) | 4.24 (3) | 2.87 (3) | 76.54 (2) | 577.60 (1) | 1.15 (1) | 2.3 |
| Simpson | 93 (6) | 0.74 ** | 8.70 * (1) | 87.48 | 19.92 (6) | 12.03 (5) | 7.89 (6) | 53.36 (1) | 591.27 (2) | 1.18 (2) | 3.6 |
| Elisa | 127 (2) | 1.45 ** | 13.67 ** (3) | 94.91 | 5.62 (2) | 4.00 (2) | 1.63 (2) | 79.94 (3) | 1284.96 (5) | 1.96 (5) | 3.0 |
| Everglades | 95 (5) | 0.84 ** | 14.93 ** (4) | 85.26 | 19.82 (5) | 11.82 (6) | 8.00 (5) | 83.93 (4) | 599.60 (3) | 1.43 (3) | 4.4 |
| Vanda | 118 (4) | 0.87 ** | 27.47 ** (5) | 78.43 | 9.23 (4) | 5.37 (4) | 3.85 (4) | 153.13 (5) | 913.96 (4) | 1.74 (4) | 4.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, M.C.; Souza, N.O.S.; Nascimento, W.M.; da Silva, G.O.; da Silva, C.R.; Suinaga, F.A. Stability Evaluation for Heat Tolerance in Lettuce: Implications and Recommendations. Plants 2024, 13, 1546. https://doi.org/10.3390/plants13111546
Pereira MC, Souza NOS, Nascimento WM, da Silva GO, da Silva CR, Suinaga FA. Stability Evaluation for Heat Tolerance in Lettuce: Implications and Recommendations. Plants. 2024; 13(11):1546. https://doi.org/10.3390/plants13111546
Chicago/Turabian StylePereira, Maryanne C., Nara O. S. Souza, Warley M. Nascimento, Giovani O. da Silva, Caroline R. da Silva, and Fabio A. Suinaga. 2024. "Stability Evaluation for Heat Tolerance in Lettuce: Implications and Recommendations" Plants 13, no. 11: 1546. https://doi.org/10.3390/plants13111546