


A Mechanistic Prediction Model of Resistance to Uprooting of Coniferous Trees in Heilongjiang Province, China

Abstract
:
1. Introduction
2. Results
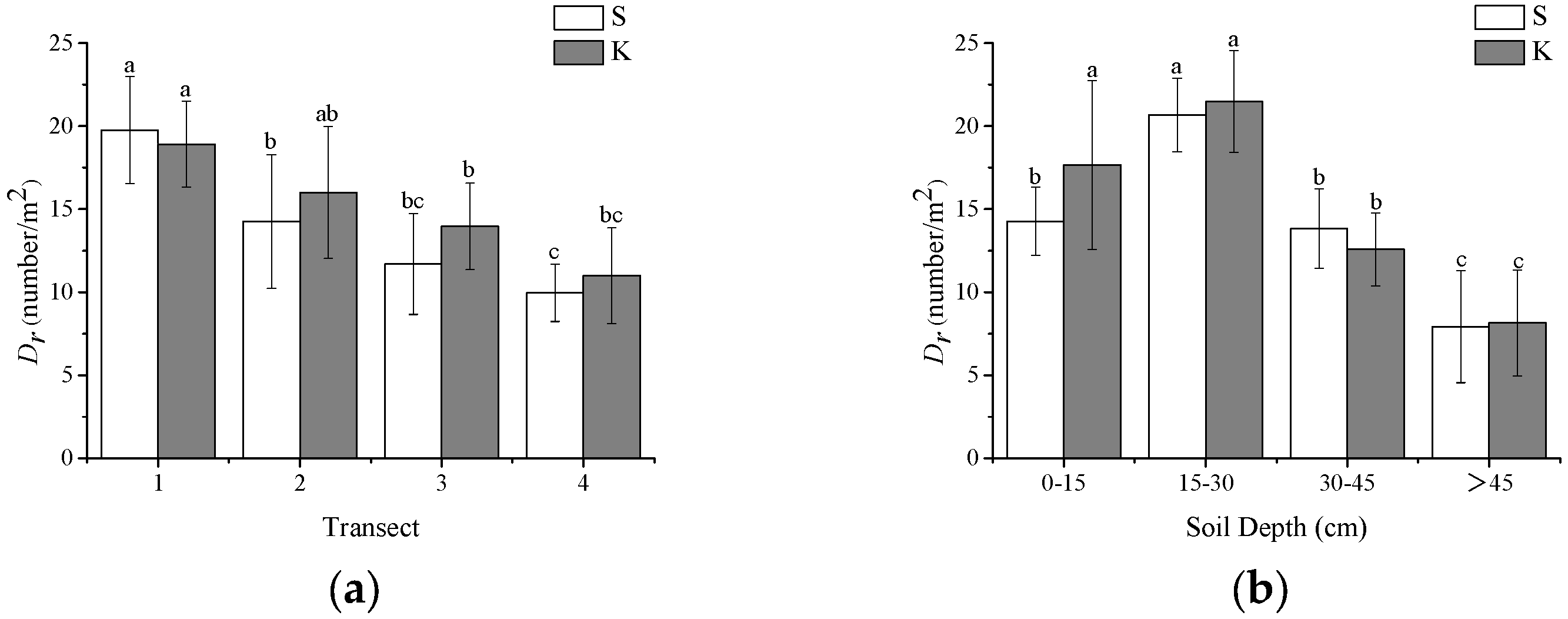
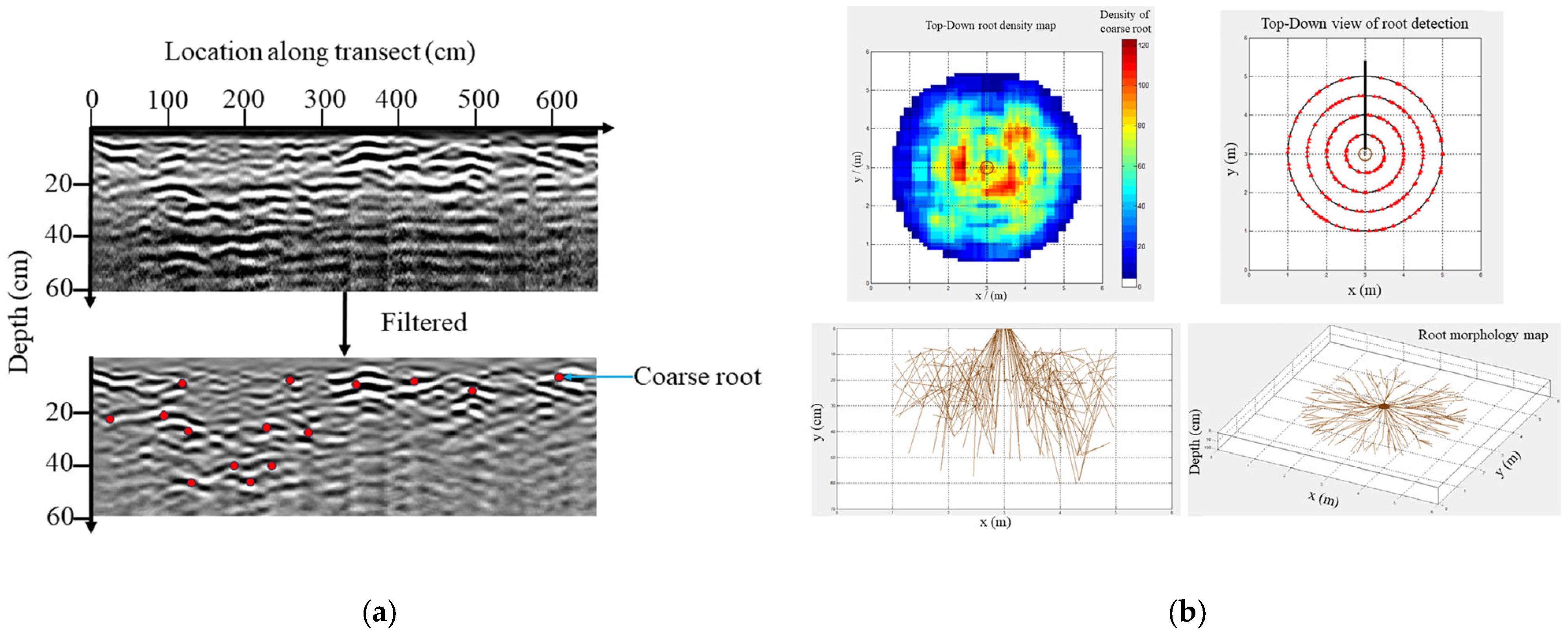
2.1. Coarse Root Distribution
2.2. Soil Properties and Root Plate Mass
2.3. Assessment of Samples in Resistance to Uprooting Using Data on Tree Morphological Characteristics
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. GPR Equipment and Data Acquisition in the Field
4.3. Soil Physical Properties
4.4. GPR Data Collection and Analusis
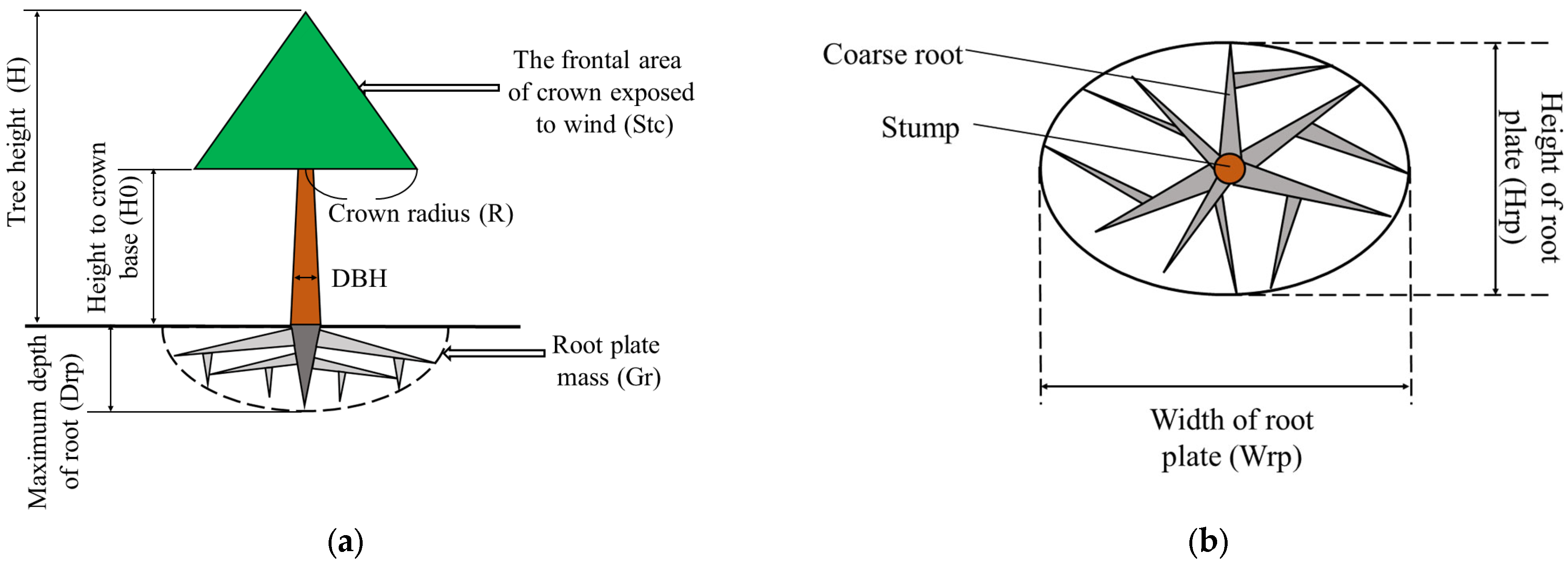
4.5. Mechanistic Model of Tree Resistance to Wind-Included Uprooting
- The tree is upright and healthy, with the root plate strong enough to support the aboveground components in windless conditions. The shape of the crown is conical. The anchorage point of the tree under wind load conditions is at the base of the stem.
- The wind load is applied horizontally and only directly to the tree crown with the assumption that the wind speed (v) is constant. The wind load is positively proportional to the frontal area of the crown (Stc) [16]. The center of gravity of the wind load is located at 2/3 of the crown length from the crown top [68,69]. The overturning moment of the wind acts on the anchorage point of the tree through the stem.
- Due to elasticity, the stem bends when wind loads are applied to the crown of the tree. However, some of the wind load can be offset by the elastic restoring force of the stem, which is positively correlated with the stiffness of the stem. The most significant factors affecting the resistive moment of the stem are tree height and DBH [55,70,71,72].
- Tree resistance to uprooting is influenced by the support provided by the root plate anchorage, which depends on a combination of four factors: the root plate mass, the strength of the windward roots and root hinge, and the soil strength at the base of the plate [26]. The mechanistic model calculates the resistive moment of the root plate based on the predicted root plate mass, as the contributions of factors other than root plate mass are very complex. The percentage contribution of root plate mass to root anchorage is constant [73,74]. Therefore, the resistive moment of the root plate can be calculated from the root plate mass, as demonstrated in the study by Peltola et al. [55]. Considering the complexity of the tree’s underground component, the root plate, which consists of coarse roots and surrounding soil, is conceptualized as a half-ellipsoid. The center of gravity of the root plate is located at 4 Drp/3 of the depth from the bottom of the root plate [75]. The mass of the root plate is mainly composed of the mass of the coarse root (Gcr) and the mass of the soil in the root plate (Gs). Gs is proportionally correlated to the bulk density of soil () and the volume of the root plate (Vr) [62,76].
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sellier, D.; Fourcaud, T. Crown structure and wood properties: Influence on tree sway and response to high winds. Am. J. Bot. 2009, 96, 885–896. [Google Scholar] [CrossRef]
- Mitchell, S. Wind as a natural disturbance agent in forests: A synthesis. For. Int. J. For. Res. 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Ge, X.; Hou, J.; Rong, B.; Wang, L.; Wang, X. Tree uprooting of coniferous broad leaved Korean pine mixed forest in Lesser Khingan Mountains, China. Chin. J. Appl. Ecol. 2016, 27, 380–386. [Google Scholar]
- Dong, L.; Zhang, L.; Li, F. A compatible system of biomass equations for three conifer species in Northeast, China. For. Ecol. Manag. 2014, 329, 306–317. [Google Scholar] [CrossRef]
- Sanginés de Cárcer, P.; Mederski, P.S.; Magagnotti, N.; Spinelli, R.; Engler, B.; Seidl, R.; Eriksson, A.; Eggers, J.; Bont, L.G.; Schweier, J. The Management Response to Wind Disturbances in European Forests. Curr. For. Rep. 2021, 7, 167–180. [Google Scholar] [CrossRef]
- Stephens, E.P. The Uprooting of Trees: A Forest Process. Soil Sci. Soc. Am. J. 1956, 20, 113–116. [Google Scholar] [CrossRef]
- Hauer, R.J.; Wang, W.; Dawson, J.O. Ice storm damage to urban trees. J. Arboric. 1993, 19, 1993. [Google Scholar] [CrossRef]
- Mou, W.P. Ice Storm Damage to Forest Tree Species in the Ridge and Valley Region of Southwestern Virginia. J. Torrey Bot. Soc. 1999, 126, 147–158. [Google Scholar]
- Rhoads, A.G.; Hamburg, S.P.; Fahey, T.J.; Siccama, T.G.; Hane, E.N.; Battles, J.; Cogbill, C.; Randall, J.; Wilson, G. Effects of an intense ice storm on the structure of a northern hardwood forest. Can. J. For. Res. 2002, 32, 1763–1775. [Google Scholar] [CrossRef]
- Ravn, H.P. Expansion of the populations of Ips typographus (L.) (Coleoptera, Scolytidae) and their local dispersal following gale disaster in Denmark. Z. Angew. Entomol. 1985, 99, 26–33. [Google Scholar] [CrossRef]
- Schroeder, L.M.; Eidmann, H.H. Attacks of bark- and wood-boring Coleoptera on snow-broken conifers over a two-year period. Scand. J. For. Res. 1993, 8, 257–265. [Google Scholar] [CrossRef]
- Gardiner, B.; Peltola, H.; Kellomäki, S. Comparison of two models for predicting the critical wind speeds required to damage coniferous trees. Ecol. Model. 2000, 129, 1–23. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Saffariha, M.; Azarnivand, H.; Tavili, A.; Khani, H.M. Investigation effects of rangeland exclosure on some soil properties in Artemisia sieberi, Stipa hohenacheriana and Salsola rigida habitats (case study: Roodshoor, Saveh, Iran). J. Biol. Environ. Sci. 2014, 4, 195–204. [Google Scholar]
- Hart, E.; Sim, K.; Kamimura, K.; Meredieu, C.; Guyon, D.; Gardiner, B. Use of machine learning techniques to model wind damage to forests. Agric. For. Meteorol. 2019, 265, 16–29. [Google Scholar] [CrossRef]
- Shibuya, M.; Koizumi, A.; Torita, H. Tree shape and resistance to uprooting: A simple model analysis. Eurasian J. For. Res. 2014, 17, 11–17. [Google Scholar]
- Blackwell, P.; Rennolls, K.; Coutts, M. A root anchorage model for shallowly rooted Sitka spruce. For. Int. J. For. Res. 1990, 63, 73–91. [Google Scholar] [CrossRef]
- Urata, T.; Shibuya, M.; Koizumi, A.; Torita, H.; Cha, J. Both stem and crown mass affect tree resistance to uprooting. J. For. Res. 2012, 17, 65–71. [Google Scholar] [CrossRef]
- Osmont, K.; Sibout, R.; Hardtke, C. Hidden Branches: Developments in Root System Architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W. The decomposition of fine and coarse roots: Their global patterns and controlling factors. Sci. Rep. 2015, 5, 9940. [Google Scholar] [CrossRef]
- Resh, S.C.; Battaglia, M.; Worledge, D.; Ladiges, S. Coarse root biomass for eucalypt plantations in Tasmania, Australia: Sources of variation and methods for assessment. Trees 2003, 17, 389–399. [Google Scholar] [CrossRef]
- Guo, L.; Lin, H.; Fan, B.; Cui, X.; Chen, J. Forward simulation of root’s ground penetrating radar signal: Simulator development and validation. Plant Soil 2013, 372, 487–505. [Google Scholar] [CrossRef]
- Song, X.; Wan, F.; Chang, X.; Zhang, J.; Liu, Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests 2019, 10, 1035. [Google Scholar] [CrossRef]
- Deans, J.D.; Ford, E.D. Modelling root structure and stability. In Tree Root Systems and Their Mycorrhizas; Atkinson, D., Bhat, K.K.S., Coutts, M.P., Mason, P.A., Read, D.J., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 189–195. [Google Scholar]
- Coutts, M.P. Components of Tree Stability in Sitka Spruce on Peaty Gley Soil. For. Int. J. For. Res. 1986, 59, 173–197. [Google Scholar] [CrossRef]
- Ray, D.; Nicoll, B.C. The effect of soil water-table depth on root-plate development and stability of Sitka spruce. For. Int. J. For. Res. 1998, 71, 169–182. [Google Scholar] [CrossRef]
- Puri, S.; Singh, V.; Bhushan, B.; Singh, S. Biomass production and distribution of roots in 3 stands of Populus deltoides. For. Ecol. Manag. 1994, 65, 135–147. [Google Scholar] [CrossRef]
- Millikin, C.S.; Bledsoe, C.S. Biomass and distribution of fine and coarse roots from blue oak (Quercus douglasii) trees in the northern Sierra Nevada foothills of California. Plant Soil 1999, 214, 27–38. [Google Scholar] [CrossRef]
- Mcivor, I.R.; Douglas, G.B.; Hurst, S.E.; Hussain, Z.; Foote, A.G. Structural root growth of young Veronese poplars on erodible slopes in the southern North Island, New Zealand. Agrofor. Syst. 2008, 72, 75–86. [Google Scholar] [CrossRef]
- Zhu, J.; Li, F.; Xu, M.; Kang, H.; Wu, X. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China: An experimental approach. Ann. For. Sci. 2008, 65, 304. [Google Scholar] [CrossRef]
- Black; Drost, D.T.; Lindstrom, T.; Reeve, J.R.; Reighard, G.L. Acomparison of root distribution patterns among Prunus rootstocks. J. Am. Pomol. Soc. 2010, 64, 52–60. [Google Scholar]
- Phillips, C.J.; Marden, M.; Suzanne, L.M. Observations of root growth of young poplar and willow planting types. N. Z. J. For. Sci. 2014, 44, 15. [Google Scholar] [CrossRef]
- Mancuso, S. (Ed.) Measuring Roots: An Updated Approach; Springer Science & Business: Boston, NY, USA, 2011. [Google Scholar]
- Hruska, J.; Čermák, J.; Šustek, S. Mapping tree root systems with ground-penetrating radar. Tree Physiol. 1999, 19, 125–130. [Google Scholar] [CrossRef]
- Jayawickreme, D.H.; Jobbágy, E.G.; Jackson, R.B. Geophysical subsurface imaging for ecological applications. New Phytol. 2014, 201, 1170–1175. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Guo, L.; Cui, X.; Chen, J.; Cao, X.; Lin, H. Ground-penetrating radar-based automatic reconstruction of three-dimensional coarse root system architecture. Plant Soil 2014, 383, 155–172. [Google Scholar] [CrossRef]
- Ohashi, M.; Ikeno, H.; Sekihara, K.; Tanikawa, T.; Dannoura, M.; Yamase, K.; Todo, C.; Tomita, T.; Hirano, Y. Reconstruction of root systems in Cryptomeria japonica using root point coordinates and diameters. Planta 2019, 249, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Butnor, J.R.; Doolittle, J.A.; Johnsen, K.H.; Samuelson, L.; Stokes, T.; Kress, L. Utility of ground-penetrating radar as a root biomass survey tool in forest systems. Soil Sci. Soc. Am. J. 2003, 67, 1607–1615. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Koteen, L.; Baldocchi, D.D. Coarse root distribution of a semi-arid oak savanna estimated with ground penetrating radar. J. Geophys. Res. Biogeosci. 2013, 118, 135–147. [Google Scholar] [CrossRef]
- Butnor, J.R.; Samuelson, L.J.; Stokes, T.A.; Johnsen, K.H.; Anderson, P.H.; González-Benecke, C.A. Surface-based GPR underestimates below-stump root biomass. Plant Soil 2016, 402, 47–62. [Google Scholar] [CrossRef]
- Simms, J.E.; McKay, S.K.; McComas, R.W.; Fischenich, J.C. In Situ Root Volume Estimation Using Ground Penetrating Radar. J. Environ. Eng. Geophys. 2017, 22, 209–221. [Google Scholar] [CrossRef]
- Zhang, T.; Song, L.; Zhu, J.; Wang, G.; Li, M.; Zheng, X.; Zhang, J. Spatial distribution of root systems of Pinus sylvestris var. mongolica trees with different ages in a semi-arid sandy region of Northeast China. For. Ecol. Manag. 2021, 483, 118776. [Google Scholar] [CrossRef]
- Wang, P.; Mommer, L.; van Ruijven, J.; Berendse, F.; Maximov, T.C.; Heijmans, M.M. Seasonal changes and vertical distribution of root standing biomass of graminoids and shrubs at a Siberian tundra site. Plant Soil 2016, 407, 55–65. [Google Scholar] [CrossRef]
- Karizumi, N. Illustrations of Tree Roots; Seibundo Shinkosha Pablishing Co., Ltd.: Tokyo, Japan, 1987. [Google Scholar]
- Xiang, S.; Zhao, X. Untersuchung Uber Die Wurzelsusteme Der Hauptanbauholzarthen in Beijing. J. Beijing For. Univ. 1981, 9–27. [Google Scholar]
- Wang, X.; Zhang, Q.; Guo, J.; Xing, T.; Wang, F.; Gao, X.; Wu, J. Study on the Coarse Root Biomass and Distribution Characteristics about Natural Larix Gmelinii Forest. For. Ecol. Manag. 2013, 5, 62. [Google Scholar] [CrossRef]
- Hamza, M.A.; Anderson, W.K. Soil compaction in cropping systems: A review of the nature, causes and possible solutions. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Yu, B.; Xie, C.; Cai, S.; Chen, Y.; Lv, Y.; Mo, Z.; Liu, T.; Yang, Z. Effects of Tree Root Density on Soil Total Porosity and Non-Capillary Porosity Using a Ground-Penetrating Tree Radar Unit in Shanghai, China. Sustainability 2018, 10, 4640. [Google Scholar] [CrossRef]
- Purushothaman, R.; Krishnamurthy, L.; Upadhyaya, H.D.; Vadez, V.; Varshney, R.K. Genotypic variation in soil water use and root distribution and their implications for drought tolerance in chickpea. Funct. Plant Biol. 2016, 44, 235–252. [Google Scholar] [CrossRef]
- Tanikawa, T.; Ikeno, H.; Todo, C.; Yamase, K.; Ohashi, M.; Okamoto, T.; Mizoguchi, T.; Nakao, K.; Kaneko, S.; Torii, A.; et al. A quantitative evaluation of soil mass held by tree roots. Trees 2021, 35, 527–541. [Google Scholar] [CrossRef]
- Yang, M.; Défossez, P.; Danjon, F.; Dupont, S.; Fourcaud, T. Which root architectural elements contribute the best to anchorage of Pinus species? Insights from in silico experiments. Plant Soil 2017, 411, 275–291. [Google Scholar] [CrossRef]
- Ilisson, T.; Metslaid, M.; Vodde, F.; Jõgiste, K.; Kurm, M. Storm disturbance in forest ecosystems in Estonia. Scand. J. For. Res. 2005, 20, 88–93. [Google Scholar]
- Kitenberga, M.; Šņepsts, G.; Vuguls, J.; Elferts, D.; Jaunslaviete, I.; Jansons, Ā. Tree-and stand-scale factors shape the probability of wind damage to birch in hemiboreal forests. Silva Fenn. 2021, 55, 10483. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical stability of Scots pine, Norway spruce and birch: An analysis of tree-pulling experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Dunham, R.A.; Cameron, A.D. Crown, stem and wood properties of wind-damaged and undamaged Sitka spruce. For. Ecol. Manag. 2000, 135, 73–81. [Google Scholar] [CrossRef]
- Fredericksen, T.; Hedden, R.; Williams, S. Testing loblolly pine wind firmness with simulated wind stress. Can. J. For. Res. 1993, 23, 1760–1765. [Google Scholar] [CrossRef]
- Cannon, J.B.; Barrett, M.E.; Peterson, C.J. The effect of species, size, failure mode, and fire-scarring on tree stability. For. Ecol. Manag. 2015, 356, 196–203. [Google Scholar] [CrossRef]
- Dupuy, L.X.; Fourcaud, T.; Lac, P.; Stokes, A. A generic 3D finite element model of tree anchorage integrating soil mechanics and real root system architecture. Am. J. Bot. 2007, 94, 1506–1514. [Google Scholar] [CrossRef]
- Danjon, F.; Fourcaud, T.; Bert, D. Root architecture and wind-firmness of mature Pinus pinaster. New Phytol. 2005, 168, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Mattheck, C.; Bethge, K.; Weber, K. The Body Language of Trees: Encyclopedia of Visual Tree Assessment, 1st ed.; Karlsruhe Institute of Technology—Campus North: Eggenstein-Leopoldshafen, Germany, 2015. [Google Scholar]
- Huang, J.; Su, Y.; Zhu, G.; Chen, R.; Wu, X. Uprooting of tree induced by typhoon: A case study of super typhoon Mangkhut in university town of Shenzhen. In E3S Web of Conferences; EDP Sciences: Les Ulis, France, 2019; p. 12012. [Google Scholar]
- Xiong, Y.; Li, Q. Soils of China; Science Press: Beijing, China, 1987. [Google Scholar]
- Zheng, B. Technical Guide for Soil Analysis; China Agriculture Press: Beijing, China, 2013. [Google Scholar]
- Burt, R. Soil Survey Field and Laboratory Methods Manual, Soil Survey Investigations Report; U.S. Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2009.
- Hirano, Y.; Dannoura, M.; Aono, K.; Igarashi, T.; Ishii, M.; Yamase, K.; Makita, N.; Kanazawa, Y. Limiting factors in the detection of tree roots using ground-penetrating radar. Plant Soil 2009, 319, 15–24. [Google Scholar] [CrossRef]
- Yan, H.; Dong, X.; Feng, G.; Zhang, S.; Mucciardi, A. Coarse root spatial distribution determined using a ground-penetrating radar technique in a subtropical evergreen broad-leaved forest, China. Sci. China Life Sci. 2013, 56, 1038–1046. [Google Scholar] [CrossRef]
- Koizumi, A. Studies on the estimation of the mechanical properties of standing trees by non-destructive bending test. Res. Bull. Coll. Exp. For. Hokkaido Univ. (Jpn.) 1987, 44, 1329–1415. [Google Scholar]
- Chiba, Y. Modelling stem breakage caused by typhoons in plantation Cryptomeria japonica forests. For. Ecol. Manag. 2000, 135, 123–131. [Google Scholar] [CrossRef]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2014, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Cucchi, V.; Meredieu, C.; Stokes, A.; Berthier, S.; Bert, D.; Najar, M.; Denis, A.; Lastennet, R. Root anchorage of inner and edge trees in stands of Maritime pine (Pinus pinaster Ait.) growing in different podzolic soil conditions. Trees 2004, 18, 460–466. [Google Scholar] [CrossRef]
- Päätalo, M.-L. Risk of Snow Damage in Unmanaged and Managed Stands of Scots Pine, Norway Spruce and Birch. Scand. J. For. Res. 2000, 15, 530–541. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S. A mechanistic model for calculating windthrow and stem breakage of Scots pines at stand edge. Silva Fenn. 1993, 27, 99–111. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Väisänen, H.; Ikonen, V. HWIND: A Mechanistic Model for Wind and Snow Damage of Scots Pine, Norway Spruce and Birch. Can. J. For. Res. 1998, 29, 647–661. [Google Scholar] [CrossRef]
- Achim, A.; Nicoll, B.C. Modelling the anchorage of shallow-rooted trees. Forestry 2009, 82, 273–284. [Google Scholar] [CrossRef]
- Nicoll, B.C.; Ray, D. Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol. 1996, 16, 891–898. [Google Scholar] [CrossRef]
- Denny, C.S.; Goodlett, J. Microrelief resulting from fallen trees. In Surficial Geology and Geomorphology of Potter County, Pennsylvania; US Government Printing Office: Washington, DC, USA, 1956; Volume 288, pp. 59–68. [Google Scholar]
- Norman, S.A.; Schaetzl, R.J.; Small, T.W. Effects of slope angle on mass movement by tree uprooting. Geomorphology 1995, 14, 19–27. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (cm) | Bulk Density (g·cm−3) * | Porosity (%) * | Water Content (%) * | Sand (%) * | Silt (%) * | Clay (%) * |
|---|---|---|---|---|---|---|
| 0~15 | 1.02 ± 0.13 | 75.35 ± 2.84 | 42.53 ± 2.41 | 34.83 ± 2.56 | 59.23 ± 2.06 | 5.94 ± 0.81 |
| 15~30 | 0.92 ± 0.15 | 79.35 ± 3.17 | 44.17 ± 2.27 | 31.36 ± 1.73 | 63.49 ± 3.18 | 7.66 ± 1.01 |
| 30~45 | 1.17 ± 0.09 | 63.93 ± 1.55 | 37.00 ± 2.84 | 28.41 ± 1.86 | 60.73 ± 1.47 | 10.86 ± 1.18 |
| >45 | 1.24 ± 0.18 | 70.6 ± 1.89 | 28.13 ± 3.66 | 30.02 ± 1.22 | 50.15 ± 1.36 | 19.83 ± 1.86 |
| Samples | Root Plate Volume (m3) | Coarse Root Mass (kg) | Soil Mass in the Root Plate (kg) | Root Plate Mass (kg) | Percentage of Coarse Root Biomass in Root Plate Mass (%) |
|---|---|---|---|---|---|
| S1 | 0.28 | 40.01 | 310.29 | 350.30 | 11.42 |
| S2 | 2.00 | 68.60 | 2175.40 | 2243.99 | 3.06 |
| S3 | 1.46 | 64.73 | 1592.00 | 1656.73 | 3.91 |
| S4 | 2.43 | 73.43 | 2652.98 | 2726.41 | 2.69 |
| S5 | 2.44 | 89.29 | 2659.25 | 2748.54 | 3.25 |
| S6 | 1.14 | 59.35 | 1246.59 | 1305.94 | 4.54 |
| S7 | 2.55 | 105.22 | 2775.02 | 2880.24 | 3.65 |
| S8 | 2.55 | 96.67 | 2775.02 | 2871.69 | 3.37 |
| S9 | 3.77 | 111.02 | 4104.98 | 4216.00 | 2.63 |
| S10 | 3.39 | 132.07 | 3695.04 | 3827.11 | 3.45 |
| S11 | 3.81 | 142.63 | 4150.04 | 4292.67 | 3.32 |
| S12 | 1.54 | 57.17 | 1675.97 | 1733.14 | 3.30 |
| S13 | 2.04 | 70.78 | 2221.96 | 2292.75 | 3.09 |
| S14 | 2.15 | 60.92 | 2344.52 | 2405.44 | 2.53 |
| S15 | 2.55 | 78.24 | 2775.02 | 2853.26 | 2.74 |
| S16 | 0.92 | 42.40 | 998.06 | 1040.45 | 4.07 |
| S17 | 0.61 | 27.98 | 662.34 | 690.33 | 4.05 |
| S18 | 2.21 | 68.78 | 2412.05 | 2480.83 | 2.77 |
| S19 | 0.28 | 17.57 | 302.04 | 319.62 | 5.50 |
| S20 | 2.29 | 70.14 | 2498.92 | 2569.06 | 2.73 |
| Mean ± S.E. | 2.02 ± 0.23 | 73.85 ± 7.16 | 2201.38 ± 250.09 | 2275.26 ± 256.79 | 3.80 ± 0.43 |
| S.D. | 1.03 | 32.02 | 1118.43 | 1148.4 | 1.94 |
| K1 | 0.46 | 32.96 | 502.35 | 535.31 | 6.16 |
| K2 | 1.43 | 40.01 | 1553.62 | 1593.64 | 2.51 |
| K3 | 3.21 | 434.70 | 3504.05 | 3938.76 | 11.04 |
| K4 | 0.30 | 23.66 | 322.81 | 346.46 | 6.83 |
| K5 | 2.08 | 181.46 | 2265.70 | 2447.17 | 7.42 |
| K6 | 4.18 | 1149.48 | 4556.20 | 5705.68 | 20.15 |
| K7 | 1.95 | 75.64 | 2125.94 | 2201.58 | 3.44 |
| K8 | 3.05 | 219.99 | 3329.27 | 3549.27 | 6.20 |
| K9 | 3.06 | 250.79 | 3338.74 | 3589.53 | 6.99 |
| K10 | 4.00 | 593.40 | 4360.00 | 4953.40 | 11.98 |
| K11 | 4.16 | 1116.47 | 4534.40 | 5650.87 | 19.76 |
| K12 | 3.45 | 497.85 | 3760.50 | 4258.35 | 11.69 |
| K13 | 0.99 | 36.76 | 1079.50 | 1116.26 | 3.29 |
| K14 | 0.64 | 34.18 | 695.34 | 729.52 | 4.69 |
| K15 | 1.45 | 40.77 | 1575.18 | 1615.94 | 2.52 |
| K16 | 0.57 | 29.37 | 624.60 | 653.97 | 4.49 |
| K17 | 1.85 | 47.16 | 2018.83 | 2065.98 | 2.28 |
| K18 | 0.66 | 35.86 | 714.07 | 749.93 | 4.78 |
| K19 | 0.67 | 40.01 | 733.72 | 773.73 | 5.17 |
| K20 | 1.76 | 41.03 | 1919.08 | 1960.11 | 2.09 |
| Mean ± S.E. | 2.00 ± 0.30 | 246.08 ± 77.98 | 2175.70 ± 324.68 | 2421.77 ± 393.11 | 7.17 ± 1.18 |
| S.D. | 1.33 | 348.73 | 1452.00 | 1758.04 | 5.30 |
| Species | DBH (cm) | Tree Height (m) | Height to Crown Base (m) | Average Crown Width (m) | Root Plate Volume (m3) | Root Plate Mass (kg) |
|---|---|---|---|---|---|---|
| Spruce | 28.60 ± 14.50 | 19.73 ± 8.18 | 11.15 ± 0.65 | 6.54 ± 1.66 | 4.24 ± 1.72 | 3407.56 ± 137.51 |
| Korean pine | 45.18 ± 11.90 | 24.30 ± 5.14 | 12.39 ± 0.62 | 7.71 ± 2.04 | 10.02 ± 2.04 | 1624.99 ± 248.14 |
| Sample | Hazard Level of Uprooting | |
|---|---|---|
| S1 | 1.18 | High |
| S2 | 1.01 | High |
| S3 | 5.32 | High |
| S4 | 0.17 | High |
| S5 | 1.51 | Low |
| S6 | 7.84 | Low |
| S7 | 0.98 | High |
| S8 | 2.73 | Low |
| S9 | 1.40 | Moderate |
| S10 | 2.82 | Low |
| S11 | 1.32 | Moderate |
| S12 | 1.65 | Low |
| S13 | 0.70 | High |
| S14 | 0.47 | High |
| S15 | 1.10 | High |
| S16 | 0.33 | High |
| S17 | 2.15 | Low |
| S18 | 0.31 | High |
| S19 | 0.70 | High |
| S20 | 0.43 | High |
| K1 | 0.83 | High |
| K2 | 0.90 | High |
| K3 | 0.71 | High |
| K4 | 1.72 | Low |
| K5 | 1.13 | High |
| K6 | 0.69 | High |
| K7 | 0.89 | High |
| K8 | 1.13 | High |
| K9 | 0.81 | High |
| K10 | 0.86 | High |
| K11 | 0.77 | High |
| K12 | 0.72 | High |
| K13 | 1.89 | Low |
| K14 | 1.59 | Low |
| K15 | 2.39 | Low |
| K16 | 1.56 | Low |
| K17 | 0.96 | High |
| K18 | 1.38 | Moderate |
| K19 | 0.69 | High |
| K20 | 1.38 | Moderate |
| Tree Number | DBH (cm) | Height to Crown Base (m) | Tree Height (m) | Crown Width | Average Crown Width (m) | |
|---|---|---|---|---|---|---|
| East-West (m) | North-South (m) | |||||
| S1 | 26.7 | 3.7 | 7.8 | 5.2 | 5.8 | 5.5 |
| S2 | 34.3 | 6.6 | 12.9 | 7.4 | 6.9 | 7.2 |
| S3 | 32.8 | 5.3 | 9.6 | 8.3 | 4.4 | 6.4 |
| S4 | 34.8 | 6.1 | 13.4 | 8.2 | 8.7 | 8.5 |
| S5 | 37.8 | 7.0 | 14.1 | 7.2 | 5.0 | 6.1 |
| S6 | 31.7 | 5.1 | 8.4 | 8.2 | 9.6 | 8.9 |
| S7 | 39.3 | 6.0 | 11.8 | 11.8 | 12.6 | 12.2 |
| S8 | 37.8 | 5.5 | 10.2 | 11.3 | 8.2 | 9.8 |
| S9 | 39.8 | 5.7 | 11.8 | 11.2 | 12.5 | 11.9 |
| S10 | 42.8 | 6.4 | 12.0 | 10.2 | 8.8 | 9.5 |
| S11 | 47.0 | 11.6 | 19.3 | 12.3 | 8.3 | 10.3 |
| S12 | 34.1 | 10.9 | 17.8 | 6.3 | 7.8 | 7.1 |
| S13 | 37.3 | 12.6 | 17.4 | 7.2 | 4.6 | 5.9 |
| S14 | 35.2 | 11.9 | 16.9 | 5.3 | 6.2 | 5.8 |
| S15 | 38.5 | 12.3 | 15.9 | 5.8 | 9.4 | 7.6 |
| S16 | 30.0 | 8.7 | 14.9 | 6.4 | 8.3 | 7.4 |
| S17 | 26.6 | 10.6 | 17.1 | 2.7 | 3.1 | 2.9 |
| S18 | 36.8 | 12.2 | 18.6 | 3.3 | 10.2 | 6.8 |
| S19 | 22.6 | 10.3 | 17.6 | 1.8 | 4.1 | 3.0 |
| S20 | 36.1 | 9.7 | 18.8 | 8.3 | 11.5 | 9.9 |
| Mean ± S.E. | 35.1 ± 1.28 | 8.41 ± 0.66 | 14.32 ± 0.81 | 7.42 ± 0.67 | 7.8 ± 0.62 | 7.64 ± 0.57 |
| S.D. | 5.71 | 2.95 | 3.63 | 2.97 | 2.77 | 2.53 |
| K1 | 25.1 | 7.3 | 16.7 | 4.1 | 3.7 | 3.9 |
| K2 | 27.2 | 12.6 | 29.1 | 5.8 | 6.7 | 6.3 |
| K3 | 64.5 | 16.5 | 35.6 | 8.6 | 8.7 | 8.7 |
| K4 | 22.3 | 7.7 | 16.6 | 6.4 | 6.4 | 6.4 |
| K5 | 46.8 | 11.8 | 24.2 | 4.8 | 8.3 | 6.6 |
| K6 | 91.7 | 18.3 | 30.9 | 12.4 | 14.6 | 13.5 |
| K7 | 34.3 | 14.6 | 29.7 | 8.6 | 8.2 | 8.4 |
| K8 | 50.5 | 16.8 | 24.8 | 7.6 | 9.8 | 8.7 |
| K9 | 52.9 | 16.2 | 33.4 | 12.8 | 13.6 | 13.2 |
| K10 | 72.0 | 14.9 | 33.8 | 9.0 | 9.6 | 9.3 |
| K11 | 90.8 | 18.9 | 35.9 | 14.7 | 15.9 | 15.3 |
| K12 | 68.2 | 23.7 | 36.2 | 11.4 | 10.2 | 10.8 |
| K13 | 26.1 | 7.2 | 16.2 | 4.1 | 5.6 | 4.9 |
| K14 | 25.5 | 8.4 | 16.8 | 7.3 | 5.6 | 6.5 |
| K15 | 27.2 | 8.9 | 14.9 | 5.8 | 6.4 | 6.1 |
| K16 | 24.2 | 9.4 | 15.6 | 2.9 | 4.7 | 3.8 |
| K17 | 28.8 | 11.4 | 17.3 | 6.9 | 5.2 | 6.1 |
| K18 | 26.0 | 9.4 | 17.2 | 9.2 | 7.9 | 8.6 |
| K19 | 26.9 | 7.1 | 17.2 | 8.6 | 7.8 | 8.2 |
| K20 | 27.2 | 7.9 | 16.5 | 8.2 | 9.6 | 8.9 |
| Mean ± S.E. | 42.9 ± 5.14 | 12.5 ± 1.07 | 23.9 ± 1.84 | 8.0 ± 0.69 | 8.4 ± 0.73 | 8.2 ± 0.69 |
| S.D. | 22.98 | 4.80 | 8.22 | 3.10 | 3.27 | 3.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, G.; Li, Y.; Wang, L.; Kan, X.; Zhang, P. A Mechanistic Prediction Model of Resistance to Uprooting of Coniferous Trees in Heilongjiang Province, China. Plants 2024, 13, 2377. https://doi.org/10.3390/plants13172377
Xie G, Li Y, Wang L, Kan X, Zhang P. A Mechanistic Prediction Model of Resistance to Uprooting of Coniferous Trees in Heilongjiang Province, China. Plants. 2024; 13(17):2377. https://doi.org/10.3390/plants13172377
Chicago/Turabian StyleXie, Guangqiang, Yaoxiang Li, Lihai Wang, Xiangcheng Kan, and Ping Zhang. 2024. "A Mechanistic Prediction Model of Resistance to Uprooting of Coniferous Trees in Heilongjiang Province, China" Plants 13, no. 17: 2377. https://doi.org/10.3390/plants13172377
APA StyleXie, G., Li, Y., Wang, L., Kan, X., & Zhang, P. (2024). A Mechanistic Prediction Model of Resistance to Uprooting of Coniferous Trees in Heilongjiang Province, China. Plants, 13(17), 2377. https://doi.org/10.3390/plants13172377