
Genome-Wide Characterization of Alfin-like Genes in Brassica napus and Functional Analyses of BnaAL02 and BnaAL28 in Response to Nitrogen and Phosphorus Deficiency

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
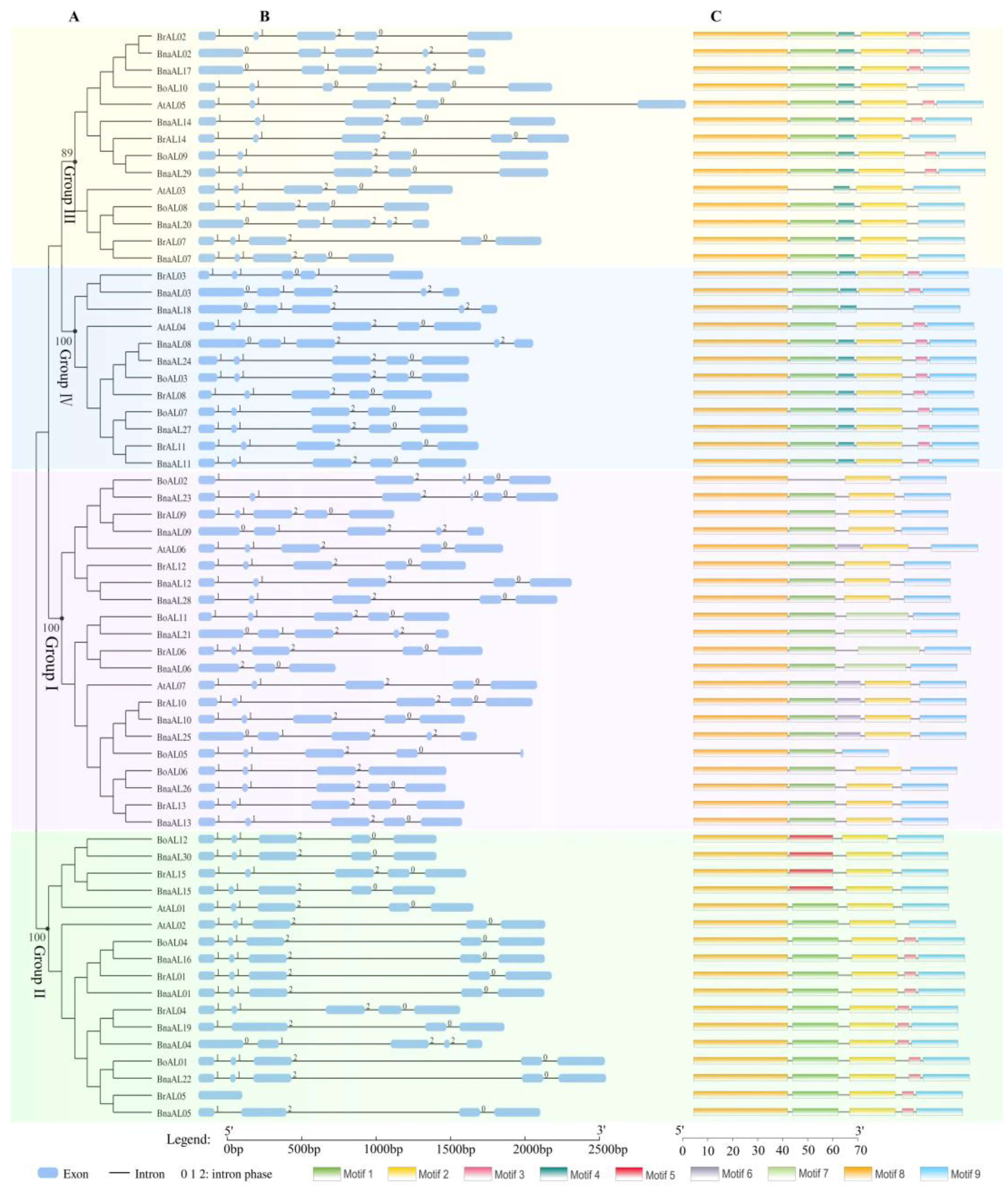
2.1. Identification and Phylogenetic Analysis of BnaALs in B. napus Genome
2.2. Gene Structural and Protein Motif Analysis of BnaALs
2.3. Allopolyploidy and Small-Scale Duplication Events Contributed to the Large Expansion of BnaALs
2.4. The Expression of BnaALs Might Be Regulated by Multiple Factors
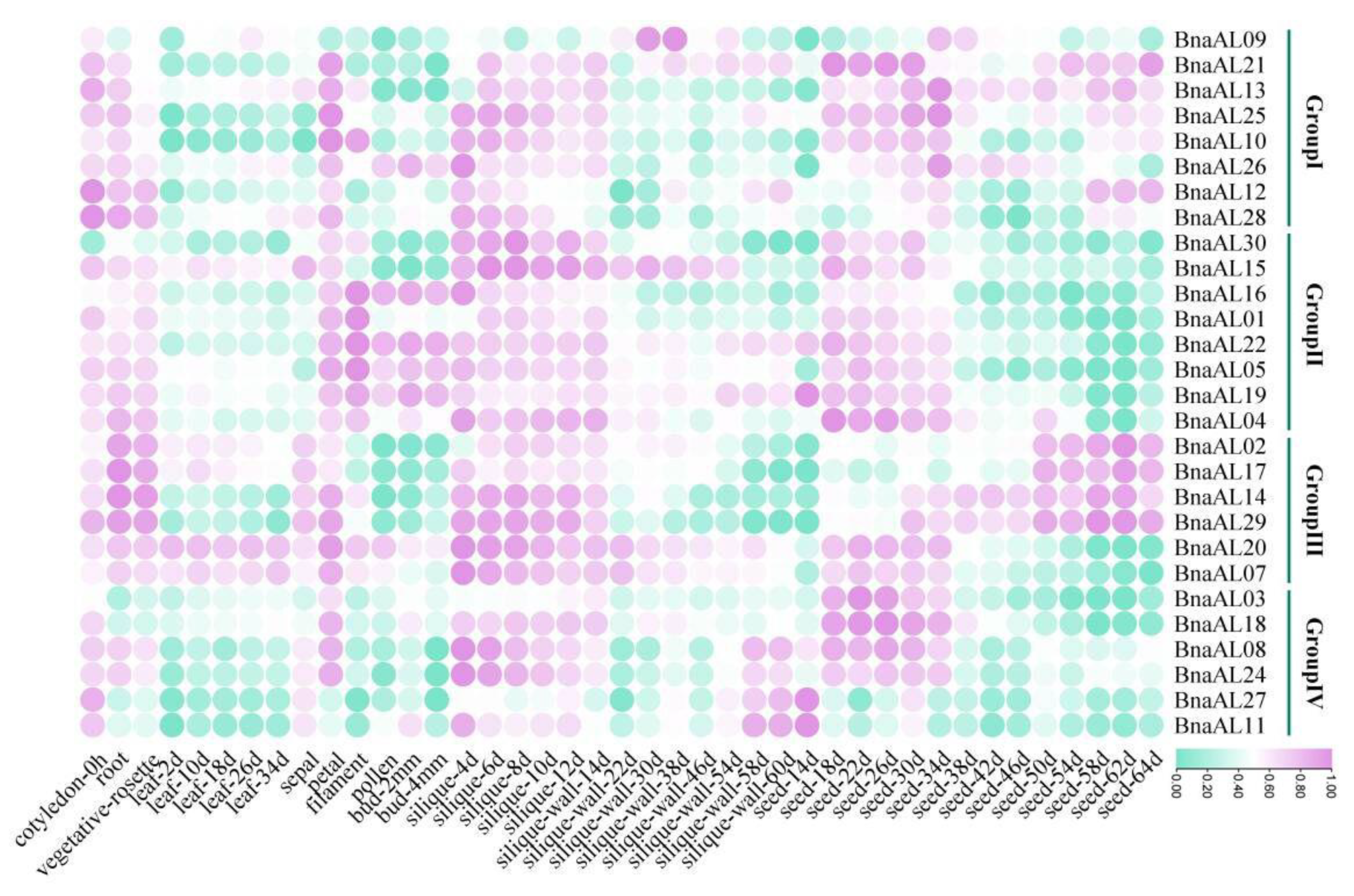
2.5. BnaALs Exhibited a Wide Spatiotemporal Expression Profiles in B. napus
2.6. BnaALs Were Inducible under LN and LP Conditions by RNA-seq and RT-qPCR Assay
2.7. BnaALs Regulated Multiple LN- and LP-Induced Roots Phenotypes in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Identification of AL Genes in B. napus and Other Plants
4.2. Phylogenetic Analysis of AL Family in B. napus
4.3. Gene Structure Analysis of BnaALs
4.4. Chromosomal Location and Collinearity Analysis of BnaALs
4.5. TF Binding Network, Cis-Elements Analysis of BnaALs
4.6. Gene Expression Analysis of BnaALs
4.7. Plant Materials and Growth Conditions
4.8. RT-qPCR Analysis of BnaALs under Low-N and Low-Pi Conditions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tao, J.-J.; Wei, W.; Pan, W.-J.; Lu, L.; Li, Q.-T.; Ma, J.-B.; Zhang, W.-K.; Ma, B.; Chen, S.-Y.; Zhang, J.-S. An Alfin-like gene from Atriplex hortensis enhances salt and drought tolerance and abscisic acid response in transgenic Arabidopsis. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, Y.; Tao, J.; Chen, H.; Li, Q.; Zhang, W.; Ma, B.; Zhang, J.; Chen, S. The Alfin-like homeodomain finger protein AL5 suppresses multiple negative factors to confer abiotic stress tolerance in Arabidopsis. Plant J. 2015, 81, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Bastola, D.R.; Pethe, V.V.; Winicov, I. Alfin1, a novel zinc-finger protein in alfalfa roots that binds to promoter elements in the salt-inducible MsPRP2 gene. Plant Mol. Biol. 1998, 38, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Huang, J.; Hao, Y.-J.; Zou, H.-F.; Wang, H.-W.; Zhao, J.-Y.; Liu, X.-Y.; Zhang, W.-K.; Ma, B.; Zhang, J.-S.; et al. Soybean GmPHD-Type Transcription Regulators Improve Stress Tolerance in Transgenic Arabidopsis Plants. PLoS ONE 2009, 4, e7209. [Google Scholar] [CrossRef]
- Lee, W.Y.; Lee, D.; Chung, W.; Kwon, C.S. Arabidopsis ING and Alfin1-like protein families localize to the nucleus and bind to H3K4me3/2 via plant homeodomain fingers. Plant J. 2009, 58, 511–524. [Google Scholar] [CrossRef]
- Winicov, I.; Bastola, D.R. Transgenic Overexpression of the Transcription Factor Alfin1 Enhances Expression of the Endogenous MsPRP2 Gene in Alfalfa and Improves Salinity Tolerance of the Plants1. Plant Physiol. 1999, 120, 473–480. [Google Scholar] [CrossRef]
- Winicov, I. Alfin1 transcription factor overexpression enhances plant root growth under normal and saline conditions and improves salt tolerance in alfalfa. Planta 2000, 210, 416–422. [Google Scholar] [CrossRef]
- Song, Y.; Gao, J.; Yang, F.; Kua, C.-S.; Liu, J.; Cannon, C.H. Molecular Evolutionary Analysis of the Alfin-Like Protein Family in Arabidopsis lyrata, Arabidopsis thaliana, and Thellungiella halophila. PLoS ONE 2013, 8, e66838. [Google Scholar] [CrossRef]
- Chandrika, N.N.P.; Sundaravelpandian, K.; Yu, S.; Schmidt, W. ALFIN-LIKE 6 is involved in root hair elongation during phosphate deficiency in Arabidopsis. New Phytol. 2013, 198, 709–720. [Google Scholar] [CrossRef]
- Jin, R.; Wang, J.; Guo, B.; Yang, T.; Hu, J.; Wang, B.; Yu, Q. Identification and Expression Analysis of the Alfin-like Gene Family in Tomato and the Role of SlAL3 in Salt and Drought Stresses. Plants 2023, 12, 2829. [Google Scholar] [CrossRef]
- Qu, L.-J.; Zhu, Y.-X. Transcription factor families in Arabidopsis: Major progress and outstanding issues for future research. Curr. Opin. Plant Biol. 2006, 9, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Kayum, A.; Park, J.-I.; Ahmed, N.U.; Jung, H.-J.; Saha, G.; Kang, J.-G.; Nou, I.-S. Characterization and stress-induced expression analysis of Alfin-like transcription factors in Brassica rapa. Mol. Genet. Genom. 2015, 290, 1299–1311. [Google Scholar] [CrossRef]
- Zhou, W.; Wu, J.; Zheng, Q.; Jiang, Y.; Zhang, M.; Zhu, S. Genome-wide identification and comparative analysis of Alfin-like transcription factors in maize. Genes Genom. 2016, 39, 261–275. [Google Scholar] [CrossRef]
- Rehman, S.U.; Sabir, I.A.; Wang, P.; Li, H.; Ahmad, Z.; Manzoor, M.A.; Zhuge, Q. Genome-wide identification of Alfin Like (AL) transcription factors and their regulatory role in abiotic stress responses in Poplar (Populus trichocarpa). Plant Stress 2023, 8, 100168. [Google Scholar] [CrossRef]
- Manzoor, M.A.; Li, G.; Xinya, W.; Wang, M.; Zhao, Y.; Sabir, I.A.; Shah, I.H.; Wang, H.; Abdullah, M.; Kim, G.-T.; et al. The Alfin-like transcription factors: Identification, characterization, and expression analysis in Pyrus bretschenedri provide insight into its divergent functions on abiotic response. Sci. Hortic. 2023, 321, 112320. [Google Scholar] [CrossRef]
- Song, J.-M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef]
- Jin, R.; Yang, H.; Muhammad, T.; Li, X.; Tuerdiyusufu, D.; Wang, B.; Wang, J. Involvement of Alfin-Like Transcription Factors in Plant Development and Stress Response. Genes 2024, 15, 184. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Luo, W.; Zheng, Y.; Guan, H.; Xie, X. Genome-wide identification and expression analysis of the PHD-finger gene family in Solanum tuberosum. PLoS ONE 2019, 14, e0226964. [Google Scholar] [CrossRef]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Liu, S.; Zhang, X.; Qian, X.; Chen, Z.; Zhao, H.; Wan, H.; Yin, N.; Li, J.; Qu, C.; et al. Genome-Wide Characterization of Alfin-like Genes in Brassica napus and Functional Analyses of BnaAL02 and BnaAL28 in Response to Nitrogen and Phosphorus Deficiency. Plants 2024, 13, 2493. https://doi.org/10.3390/plants13172493
Wu Z, Liu S, Zhang X, Qian X, Chen Z, Zhao H, Wan H, Yin N, Li J, Qu C, et al. Genome-Wide Characterization of Alfin-like Genes in Brassica napus and Functional Analyses of BnaAL02 and BnaAL28 in Response to Nitrogen and Phosphorus Deficiency. Plants. 2024; 13(17):2493. https://doi.org/10.3390/plants13172493
Chicago/Turabian StyleWu, Zexuan, Shiying Liu, Xinyun Zhang, Xingzhi Qian, Zhuo Chen, Huiyan Zhao, Huafang Wan, Nengwen Yin, Jiana Li, Cunmin Qu, and et al. 2024. "Genome-Wide Characterization of Alfin-like Genes in Brassica napus and Functional Analyses of BnaAL02 and BnaAL28 in Response to Nitrogen and Phosphorus Deficiency" Plants 13, no. 17: 2493. https://doi.org/10.3390/plants13172493