
Photosynthesis, Chlorophyll Fluorescence, and Hormone Regulation in Tomato Exposed to Mechanical Wounding

Abstract
1. Introduction
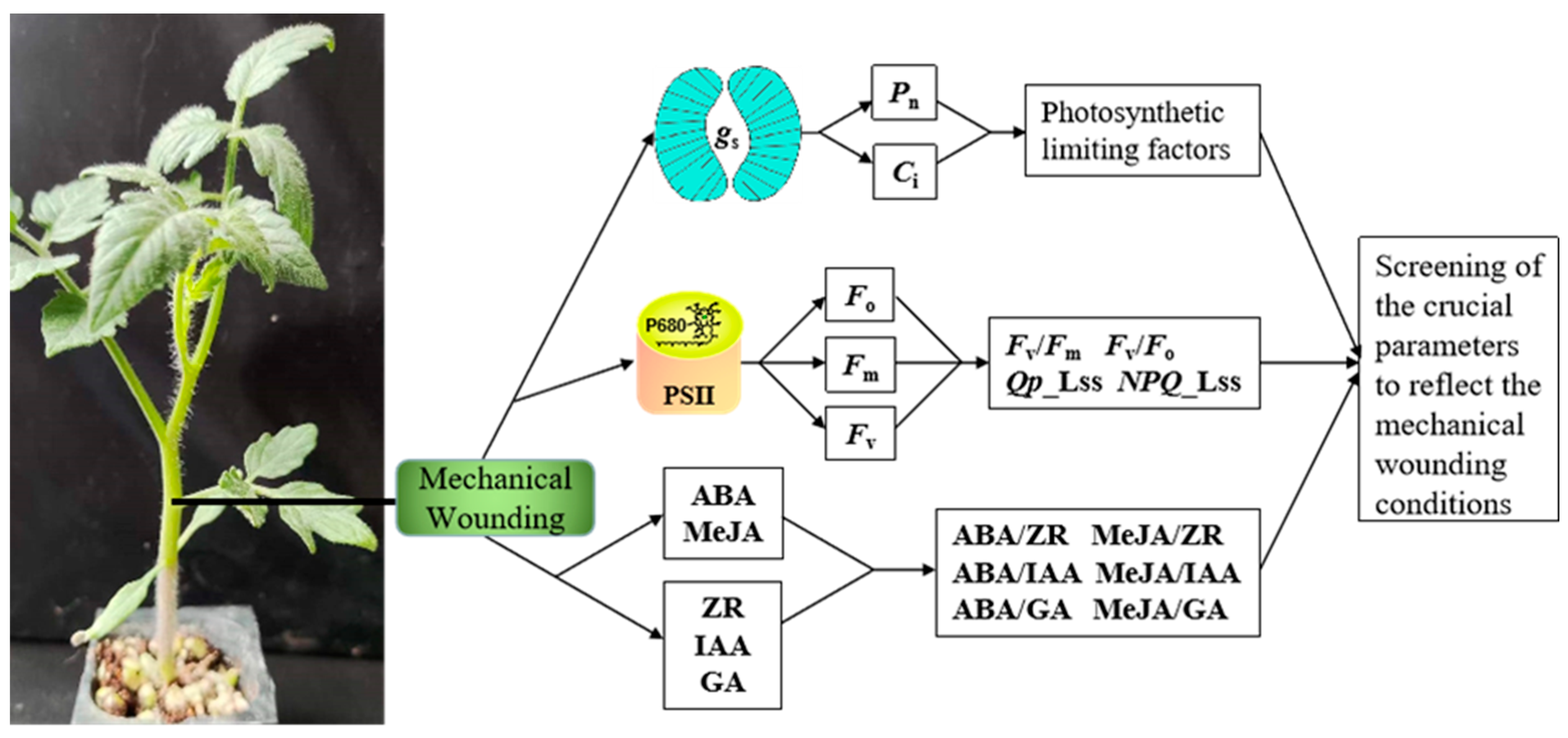
2. Materials and Methods
2.1. Plant Cultivation
2.2. Compression Treatment
2.3. Photosynthesis Measurements
2.4. Fluorescence Measurements
2.5. Phytohormone Assay
2.6. Statistical Analysis
3. Results
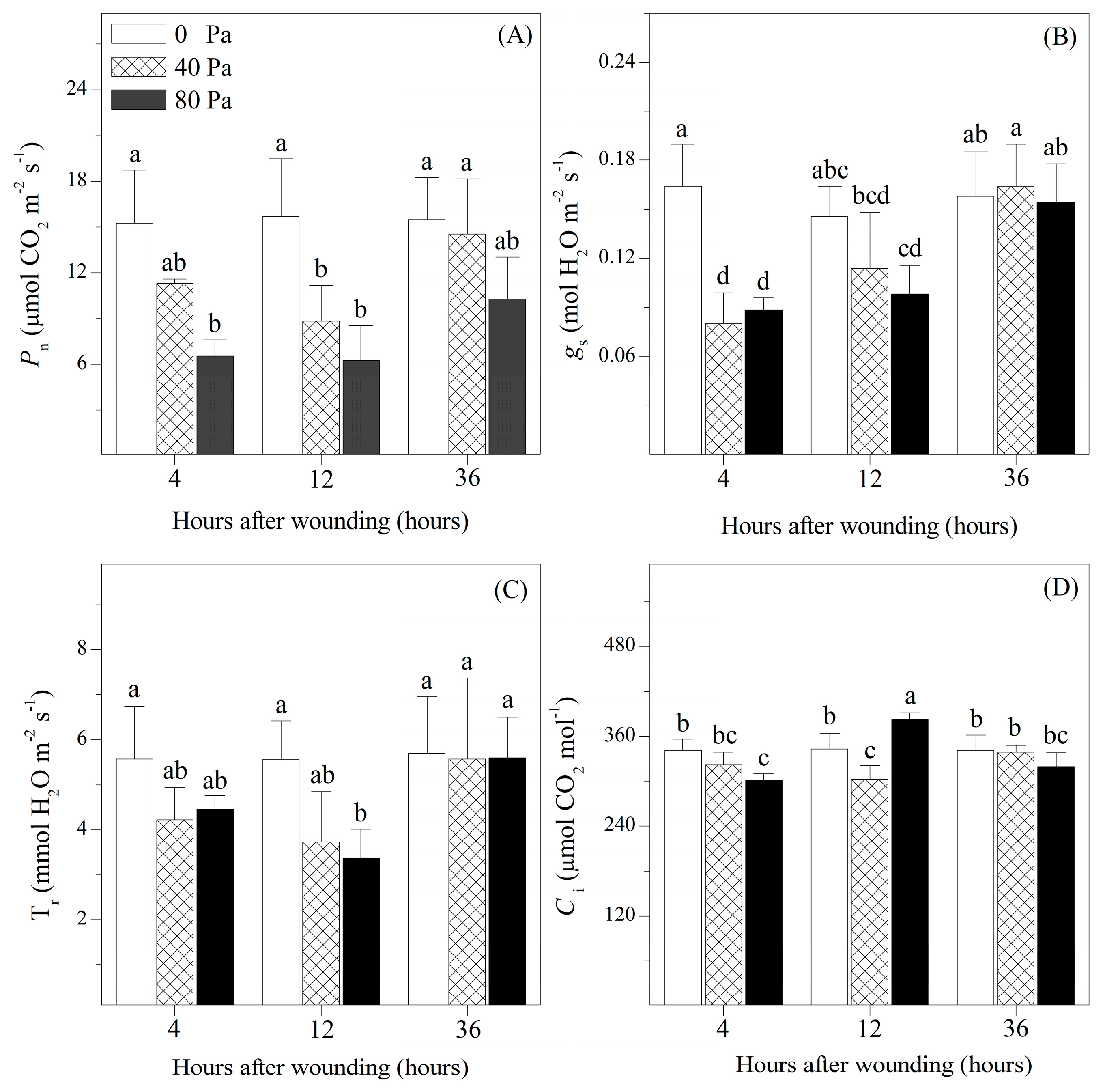
3.1. Photosynthetic Gas Exchange
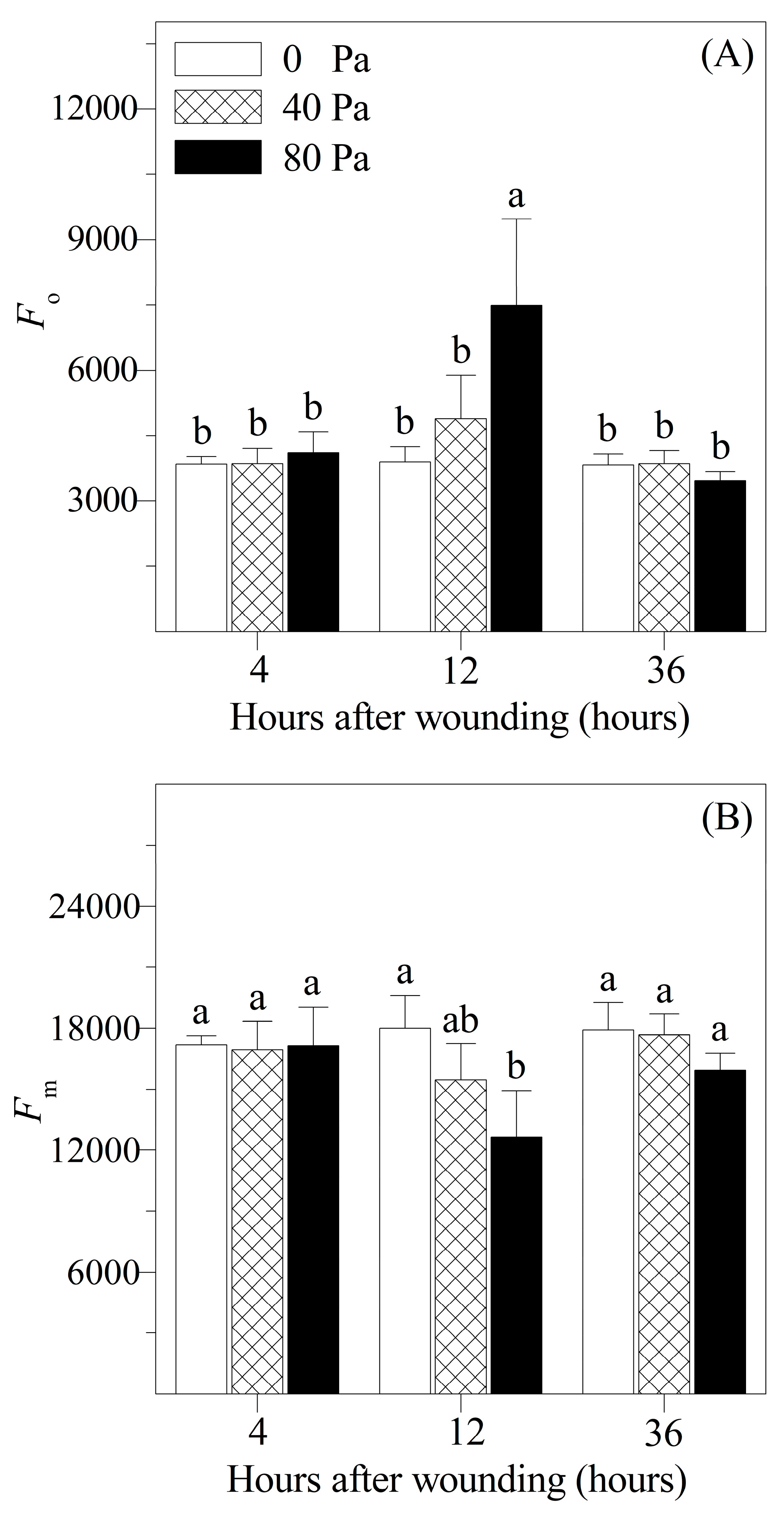
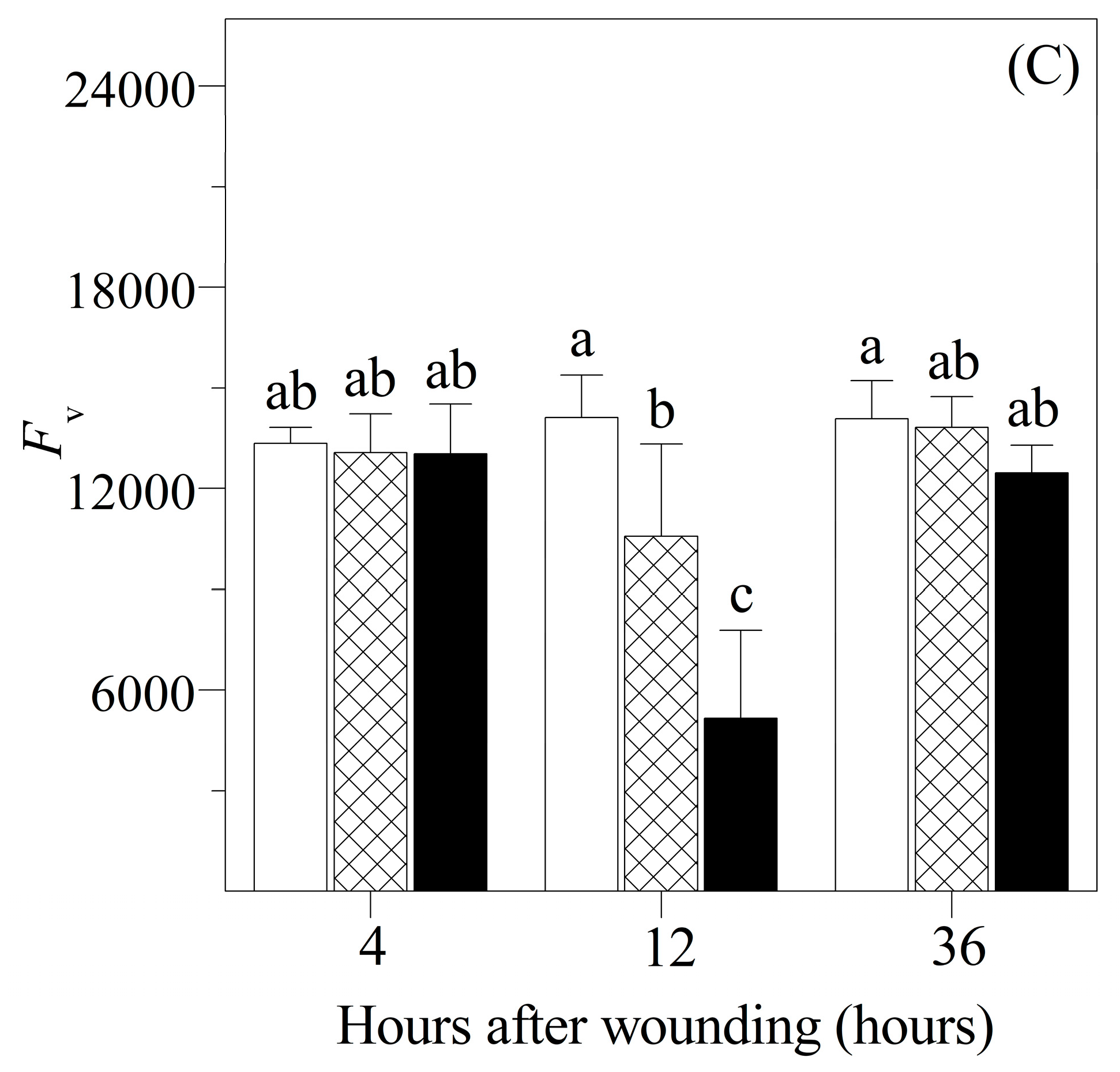
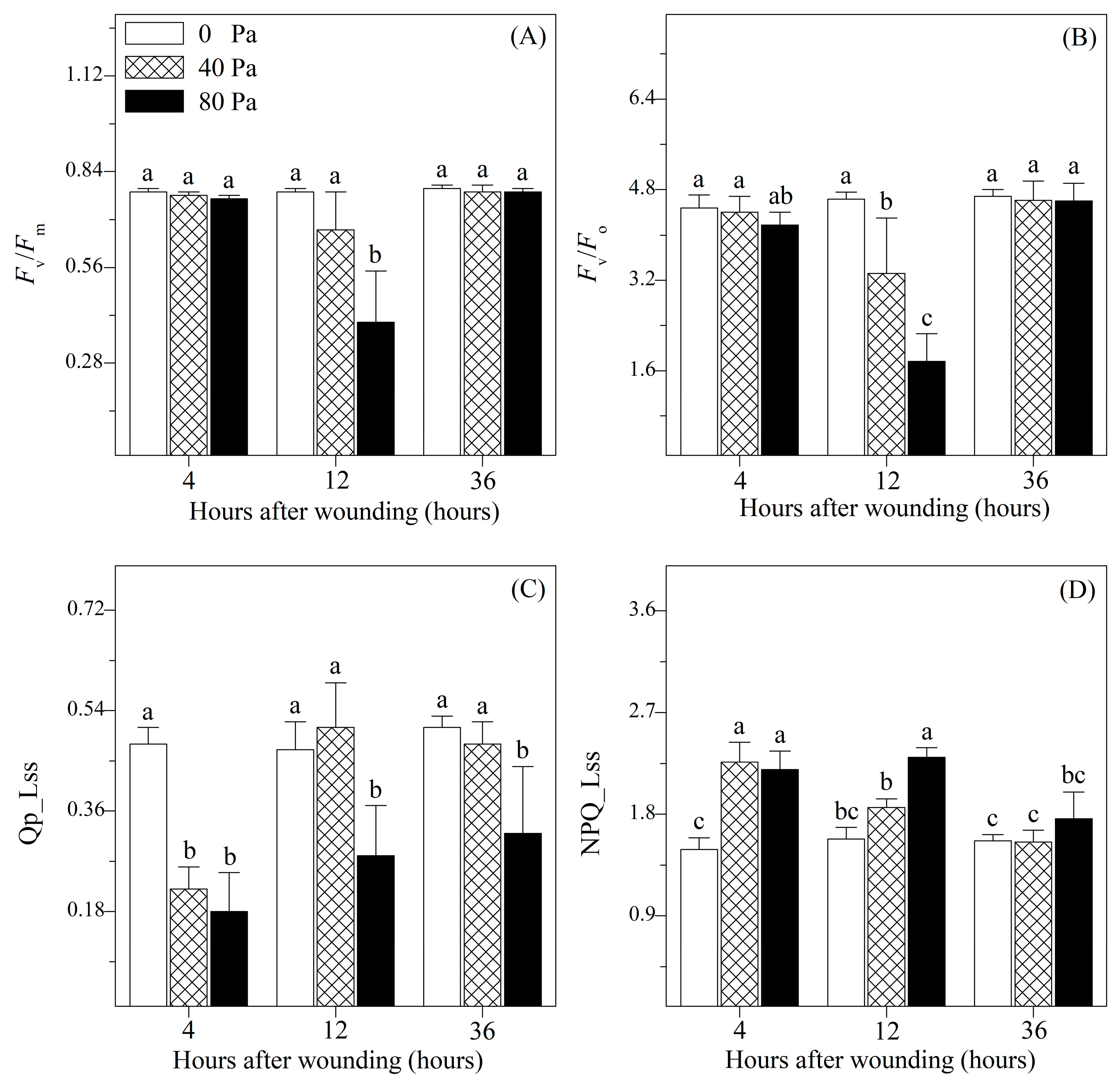
3.2. Chlorophyll Fluorescence
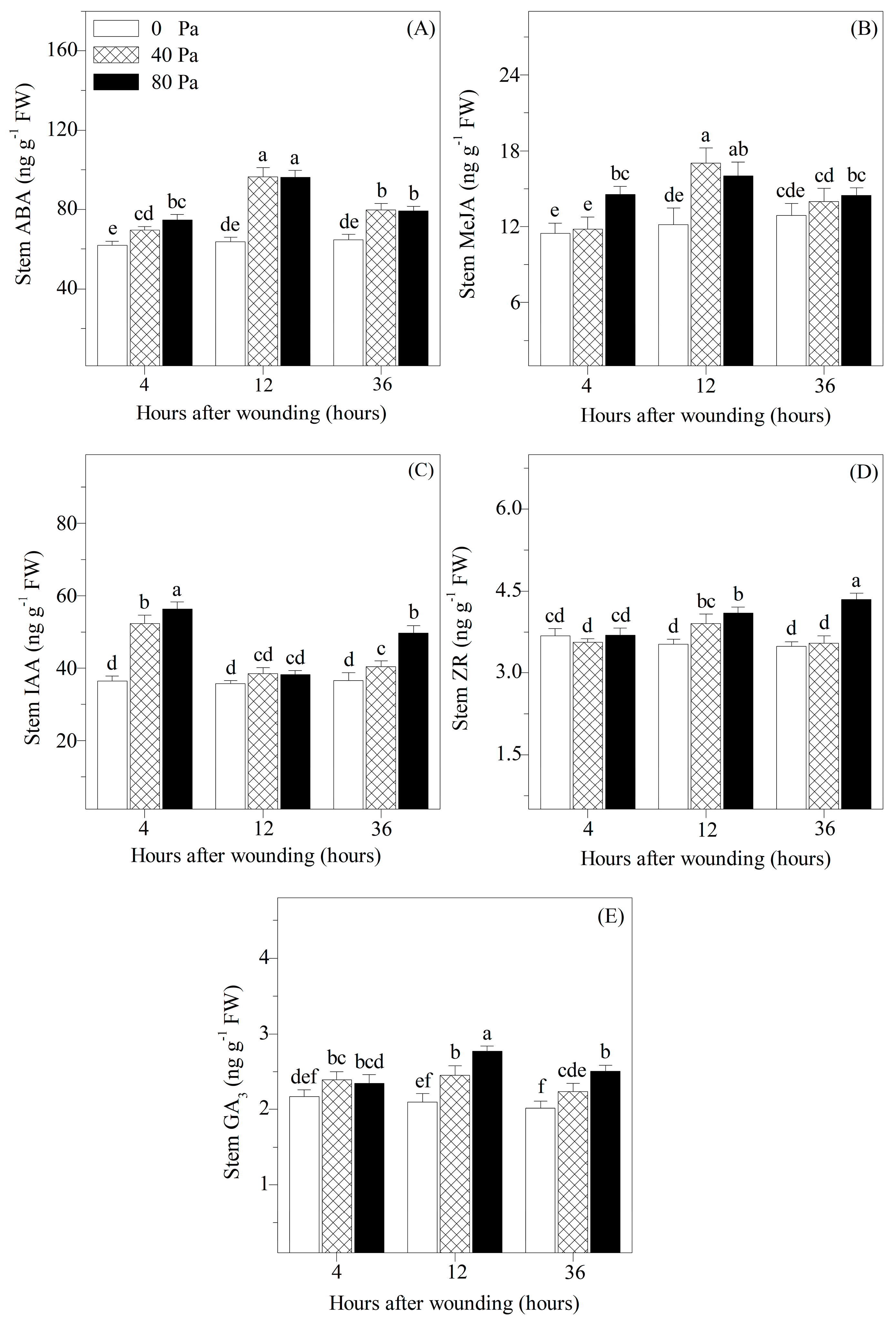
3.3. Concentrations of Stress-Responsive and Growth-Promoting Hormones in Stems
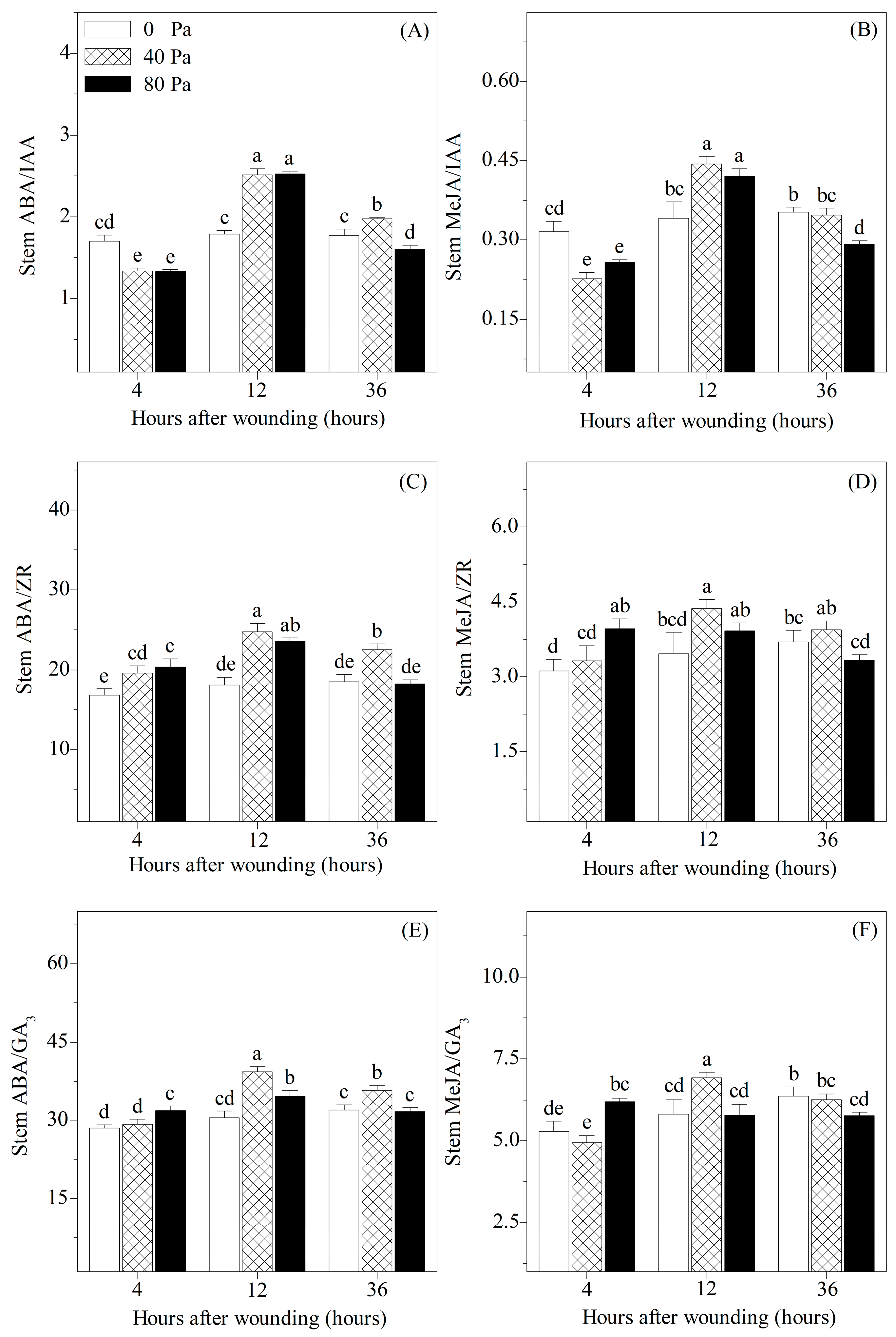
3.4. Ratios of Stress-Responsive Hormones to Growth-Promoting Hormones in the Stems
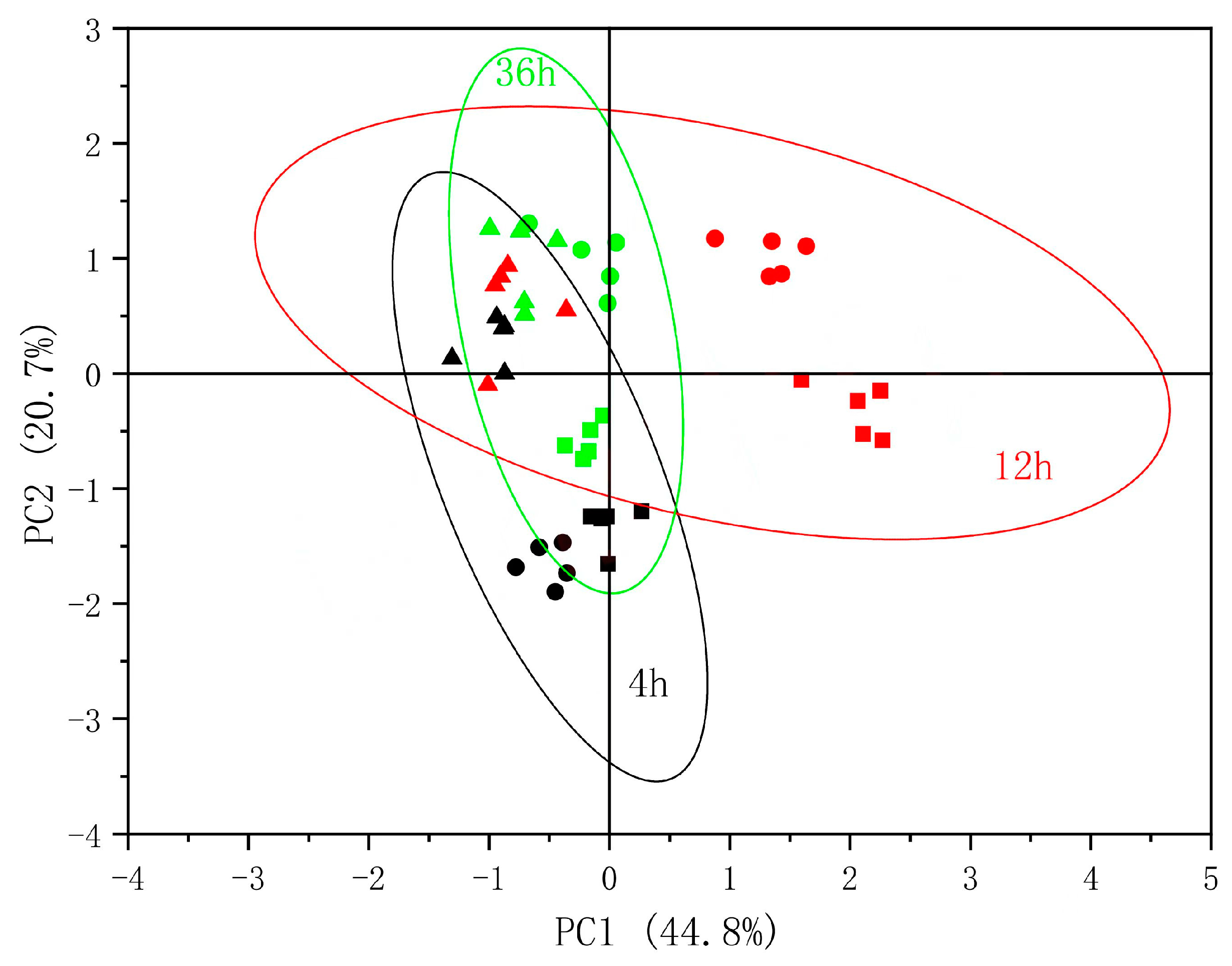
3.5. PCA of Physiological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hu, S.; Hu, M.; Zhang, W. Design and experiment of flexible clamping device for pepper plug seedlings. Adv. Mech. Eng. 2022, 14, 6. [Google Scholar]
- León, J.; Rojo, E.; Sánchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lukaszuk, E.; Rys, M.; Możdżeń, K.; Stawoska, I.; Skoczowski, A.; Ciereszko, I. Photosynthesis and sucrose metabolism in leaves of Arabidopsis thaliana aos, ein4 and rcd1 mutants as affected by wounding. Acta Physiol. Plant. 2017, 39, 17. [Google Scholar] [CrossRef]
- Wang, S.J.; Lan, Y.C.; Chen, S.F.; Chen, Y.M.; Yeh, K.W. Wound-response regulation of the sweet potato sporamin gene promoter region. Plant Mol. Biol. 2002, 48, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, C.Q. Mechanism of wound-induced defense response and signal transduction in plants. Chin. Bull. Bot. 2013, 48, 461–469. [Google Scholar]
- Lulai, E.C.; Suttle, J.C.; Pederson, S.M. Regulatory involvement of abscisic acid in potato tuber wound-healing. J. Exp. Bot. 2008, 59, 1175–1186. [Google Scholar] [CrossRef]
- Koo, A.J.K.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef]
- Howe, G.A. Jasmonates as signals in the wound response. J. Plant Growth Regul. 2004, 23, 223–237. [Google Scholar] [CrossRef]
- Lulai, E.C.; Suttle, J.C.; Olson, L.L.; Neubauer, J.D.; Campbell, L.G.; Campbell, M.A. Wounding induces changes in cytokinin and auxin content in potato tuber, but does not induce formation of gibberellins. J. Plant Physiol. 2016, 191, 22–28. [Google Scholar] [CrossRef]
- Brenya, E.; Chen, Z.H.; Tissue, D.; Papanicolaou, A.; Cazzonelli, C.I. Prior exposure of Arabidopsis seedlings to mechanical stress heightens jasmonic acid-mediated defense against necrotrophic pathogens. BMC Plant Biol. 2020, 20, 548. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, N.; Yang, R.; Zhang, X.; Wang, Y.; Li, Y.; Prusky, D.; Bi, Y.; Han, Y. Essential role of ABA signaling and related transcription factors in phenolic acid and lignin synthesis during muskmelon wound healing. Front. Plant Sci. 2024, 15, 1404477. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Wu, L.; Filardo, F.; Yang, X.; Zhao, X.; Fu, D. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant. 2017, 39, 125. [Google Scholar] [CrossRef]
- Sarabi, B.; Fresneau, C.; Ghaderi, N.; Bolandnazar, S.; Streb, P.; Badeck, F.; Citerne, S.; Tangama, M.; David, A.; Ghashghaie, J. Stomatal and non-stomatal limitations are responsible in down-regulation of photosynthesis in melon plants grown under the saline condition: Application of carbon isotope discrimination as a reliable proxy. Plant Physiol. Bioch. 2019, 141, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.; Rashkov, G.; Borisova, P.; Apostolova, E. Sensitivity of the Photosynthetic apparatus in maize and sorghum under different drought levels. Plants 2023, 12, 1863. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Hu, X.; Li, F. Leaf photosynthesis, chlorophyll fluorescence, ion content and free amino acids in Caragana korshinskii Kom exposed to NaCl stress. Acta Physiol. Plant. 2012, 34, 2285–2295. [Google Scholar] [CrossRef]
- Vats, S.; Bansal, R.; Rana, N.; Kumawat, S.; Bhatt, V.; Jadhav, P.; Kale, V.; Sathe, A.; Sonah, H.; Jugdaohsingh, R.; et al. Unexplored nutritive potential of tomato to combat global malnutrition. Crit. Rev. Food Sci. Nutr. 2022, 62, 1003–1034. [Google Scholar] [CrossRef]
- Prasad, R.; Lisiecka, J.; Antala, M.; Rastogi, A. Influence of different spent mushroom substrates on yield, morphological and photosynthetic parameters of strawberry (Fragaria × ananassa Duch.). Agronomy 2021, 11, 2086. [Google Scholar] [CrossRef]
- He, Z. A Laboratory Guide to Chemical Control Technology on Field Crop; Beijing Agricultural University Press: Beijing, China, 1993. [Google Scholar]
- Kumar, K.; Giri, A.; Vivek, P.; Kalaiyarasan, T.; Kumar, B. Effects of fluoride on respiration and photosynthesis in plants: An overview. Ann. Environ. Sci. Toxicol. 2017, 2, 43–47. [Google Scholar] [CrossRef]
- Tartachnyk, I.; Blanke, M.M. Effect of mechanically-simulated hail on photosynthesis, dark respiration and transpiration of apple leaves. Environ. Exp. Bot. 2002, 48, 169–175. [Google Scholar] [CrossRef]
- Yan, H.; Liu, X.; Ding, H.; Dai, Z.; Niu, X.; Zhao, L. Hormonal balance, photosynthesis, and redox reactions in the leaves of Caragana korshinskii Kom. under water deficit. Plants 2023, 12, 2076. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- de Souza, G.A.R.; de Andrade Neves, D.; Ruas, K.F.; Baroni, D.F.; dos Santos, K.D.; de Paula Bernado, W.; dos Santos Nani de Miranda, R.M.; de Almeida, C.M.; Santos, A.R.; Rodrigues, W.P.; et al. Supra-optimal temperatures induce photochemical leaf damage and reduce photosynthetic O2 evolution in Carica papaya L. Environ. Exp. Bot. 2022, 203, 105051. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Fang, X.; Wang, K.; Sun, X.; Wang, Y.; Zheng, P.; Shi, F. Characteristics of chlorophyll fluorescence in ten garden shrub species under flooding stress. Biologia 2022, 77, 339–350. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagôa, A.M.M.A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and mechanism of jasmonic acid in plant responses to abiotic and biotic stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Lee, G.I.; Howe, G.A. Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato. Proc. Natl. Acad. Sci. USA 2002, 99, 6416–6421. [Google Scholar] [CrossRef]
- Khan, A.L.; Al-Harrasi, A.; Shahzad, R.; Imran, Q.M.; Yun, B.W.; Kim, Y.H.; Kang, S.M.; Al-Rawahi, A.; Lee, I.J. Regulation of endogenous phytohormones and essential metabolites in frankincense-producing Boswellia sacra under wounding stress. Acta Physiol. Plant. 2018, 40, 113. [Google Scholar] [CrossRef]
- Pan, X.; Welti, R.; Wang, X. Simultaneous quantification of major phytohormones and related compounds in crude plant extracts by liquid chromatography-electrospray tandem mass spectrometry. Phytochemistry 2008, 69, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Battal, P.; Erez, M.E.; Turker, M.; Berber, I. Molecular and physiological changes in maize (Zea mays) induced by exogenous NAA, ABA and MeJa during cold stress. Ann. Bot. Fennici. 2008, 45, 173–185. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Front. Plant Sci. 2018, 46, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.L.; Chen, Q.; Chen, Q.J.; Jiang, M.; Gao, W.; Qu, Y.Y. Screening of key drought tolerance indices for cotton at the flowering and boll setting stage using the dimension reduction method. Front. Plant Sci. 2021, 12, 619926. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Pn | −0.20 | 0.22 | −0.17 | −0.02 |
| Tr | −0.20 | 0.15 | −0.08 | 0.35 |
| gs | −0.14 | 0.30 | −0.14 | 0.39 |
| Ci | 0.06 | 0.10 | −0.39 | −0.06 |
| Fo | 0.24 | −0.01 | −0.28 | −0.21 |
| Fm | −0.24 | 0.07 | 0.19 | −0.24 |
| Fv | −0.27 | 0.05 | 0.26 | −0.07 |
| Fv/Fm | −0.26 | 0.03 | 0.29 | 0.09 |
| Fv/Fo | −0.28 | 0.02 | 0.23 | 0.08 |
| Qp_Lss | −0.07 | 0.41 | 0.00 | −0.02 |
| NPQ_Lss | 0.17 | −0.33 | 0.02 | −0.20 |
| ABA | 0.30 | 0.03 | 0.12 | 0.14 |
| MeJA | 0.27 | 0.06 | 0.24 | 0.21 |
| IAA | −0.01 | −0.38 | 0.27 | 0.17 |
| ZR | 0.16 | −0.09 | −0.04 | 0.60 |
| GA3 | 0.26 | −0.19 | −0.03 | 0.26 |
| ABA/IAA | 0.24 | 0.27 | −0.09 | −0.05 |
| ABA/ZR | 0.27 | 0.09 | 0.19 | −0.16 |
| ABA/GA3 | 0.22 | 0.23 | 0.24 | −0.03 |
| MeJA/IAA | 0.21 | 0.33 | −0.04 | −0.03 |
| MeJA/ZR | 0.21 | 0.14 | 0.32 | −0.12 |
| MeJA/GA3 | 0.12 | 0.28 | 0.36 | 0.02 |
| Proportion of variation (%) | 44.77 | 20.73 | 13.35 | 6.27 |
| Cumulative proportions of variation (%) | 44.77 | 65.50 | 78.85 | 85.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Fu, K.; Li, J.; Li, M.; Li, S.; Dai, Z.; Jin, X. Photosynthesis, Chlorophyll Fluorescence, and Hormone Regulation in Tomato Exposed to Mechanical Wounding. Plants 2024, 13, 2594. https://doi.org/10.3390/plants13182594
Yan H, Fu K, Li J, Li M, Li S, Dai Z, Jin X. Photosynthesis, Chlorophyll Fluorescence, and Hormone Regulation in Tomato Exposed to Mechanical Wounding. Plants. 2024; 13(18):2594. https://doi.org/10.3390/plants13182594
Chicago/Turabian StyleYan, Hui, Kai Fu, Jiajia Li, Mingyong Li, Shaofan Li, Zhiguang Dai, and Xin Jin. 2024. "Photosynthesis, Chlorophyll Fluorescence, and Hormone Regulation in Tomato Exposed to Mechanical Wounding" Plants 13, no. 18: 2594. https://doi.org/10.3390/plants13182594
APA StyleYan, H., Fu, K., Li, J., Li, M., Li, S., Dai, Z., & Jin, X. (2024). Photosynthesis, Chlorophyll Fluorescence, and Hormone Regulation in Tomato Exposed to Mechanical Wounding. Plants, 13(18), 2594. https://doi.org/10.3390/plants13182594