
Efficient Plantlet Regeneration from Branches in Mangifera indica L.

 ,
,
Abstract
1. Introduction
2. Results
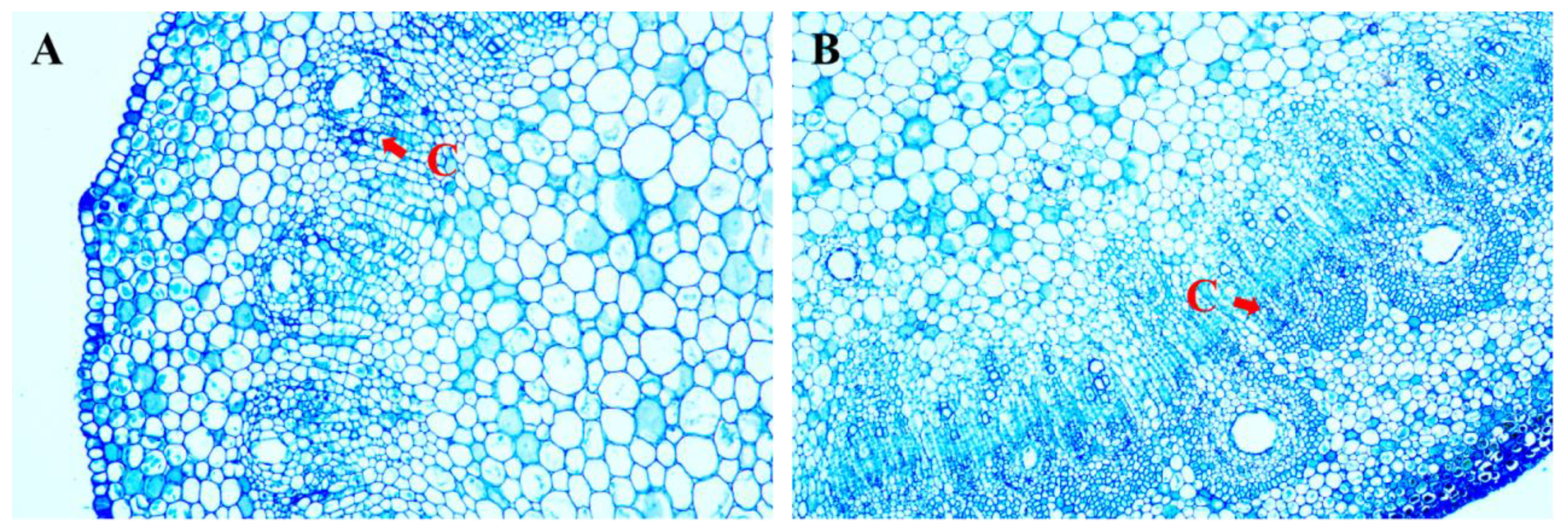
2.1. Adventitious Shoot Induction from Branches of Mangifera indica L.
2.2. Effect of PGRs on Adventitious Shoot Induction
2.3. Effect of Basic Culture Medium on Induction of Adventitious Shoots
2.4. Effects of 6-BA on Elongation and Growth Status
2.5. Effect of IBA on Rooting of Mangifera indica L.
2.6. Acclimatization
2.7. Genetic Fidelity Assessment
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Culture Conditions
4.2. Preparation of Sterile Materials
4.3. Induction of Adventitious Shoots from Branches
4.4. Elongation of Adventitious Shoots
4.5. Induction of Roots and Acclimatization
4.6. Genetic Homogeneity Analysis
4.7. Statistic and Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.J.; Li, F.; Yuan, N.; Zhang, H. Research Progress of Mango Tissue Culture. Fujian For. Sci. Technol. 2021, 48, 121–124. [Google Scholar]
- Hu, Y.; Zhang, D.S.; Liu, K.D. Developing History and Influence Factors of the Mango Industry in China. Chin. J. Agric. Resour. Reg. Plan. 2015, 36, 53–59. [Google Scholar]
- Duan, Y.J.; Yang, Y.J.; Meng, F.X.; Liu, H.G. Research progress on mango (Mangifera indica L.) seedling propagation technology. Subtrop. Agric. Res. 2017, 13, 205–210. [Google Scholar]
- Zhong, Z. How to cultivate mango grafted rootstock adventitious shoots. Rural. Pract. Technol. 2007, 1, 46. [Google Scholar]
- Krishna, H.; Singh, S.K. Biotechnological advances in mango (Mangifera indica L.) and their future implication in crop improvement—A review. Biotechnol. Adv. 2007, 25, 223–243. [Google Scholar] [CrossRef]
- Pan, Q.C. Mango grafting technology and seedling management. Chin. Hortic. Dig. 2010, 26, 132–133. [Google Scholar]
- Ou, Z.Y.; Zhao, D.M.; He, D.G. Meteorological conditions and production management measures for mango grafting survival. Meteorol. Sci. Technol. 2010, 38, 133–135. [Google Scholar]
- Ai, M. Different grafting periods and survival of mangoes. Yunnan Trop. Sci. Technol. 1990, 28, 14. [Google Scholar]
- Liu, Y.; Jiang, J.Y. Brief report on the rapid seedling cultivation experiment of mango seedlings by cutting and grafting. Fujian Fruit. Tree 1996, 1, 24. [Google Scholar]
- Tian, J.W.; Yang, C.Z.; Chen, M.X.; Huang, Y.J.; Cao, Y.C.; Chen, D.J.; Zhu, X.K.; Zhang, Q.Y. Preliminary report on mango cutting seedling cultivation experiment. For. Sci. Technol. Commun. 1991, 7, 20–21. [Google Scholar]
- Nan, S.; Asla, M.M. Effect of different concentrations of indole butyric acid (IBA) and age of shoot on air layering of mango (Mangifera indica Linn). J. Res. 2003, 14, 193–198. [Google Scholar]
- Baghel, B.S. Response of air layering of mango to coloured polywarappers. Indian J. Hortic. 1999, 56, 133–134. [Google Scholar]
- Litz, R.E.; Knight, R.L.; Gazit, S.B. Somatic embryos from culture ovules of polyembryonic Mangifera indica L. Plant Cell Rep. 1982, 1, 264–266. [Google Scholar] [CrossRef] [PubMed]
- DeWald, S.G.; Litz, R.E.; Moore, G.A. Maturation and germination of mango somatic embryos. J. Am. Soc. Hortic. Sci. 1989, 114, 837–841. [Google Scholar] [CrossRef]
- Raghuvanshi, S.S.; Srivastava, A. Plant regeneration of Mangifera indica using liquid shaker culture to reduce phenolic exudation. Plant Cell Tissue Organ. Cult. 1995, 41, 83–85. [Google Scholar] [CrossRef]
- Wu, Y. A Study on Somatic Embryogenesis and Cryopreservation of Embryogenic Culture of Mango (Mangifera indica L.); Sun Yat-sen University: Guangzhou, China, 2005; pp. 1–24. [Google Scholar]
- Xiao, J. Regulation of In Vitro Morphogenesis of Mangifera indica L. and Cloning of Genes Related to Adventitious Root Formation; Sun Yat-sen University: Guangzhou, China, 2003; pp. 1–29. [Google Scholar]
- Conde, F.; Carmona-Martin, E.; Hormaza, J.I.; Petri, C. In vitro establishment and micropropagation of mango (Mangifera indica L.) from cotyledonary nodes. Vitr. Cell. Dev. Biol.-Plant 2023, 59, 197–208. [Google Scholar] [CrossRef]
- Petri, C.; Litz, R.E.; Singh, S.K.; Hormaza, J.I. In vitro culture and genetic transformation in mango. In The Mango Genome; Kole, C., Ed.; Springer Nature: Cham, Switzerland, 2021; pp. 131–153. [Google Scholar]
- Zhang, Y.Y.; Wei, J.Y.; Li, K.M. Progress in the Research of Tissue Culture of Mango. Chin. J. Trop. Crops 2010, 31, 325–331. [Google Scholar]
- Bettoni, J.C.; Bonnart, R.; Volk, G.M. Challenges in implementing plant shoot tip cryopreservation technologies. Plant Cell Tiss. Organ. Cult. 2021, 144, 21–34. [Google Scholar] [CrossRef]
- Solangi, N.; Jatoi, M.A.; Mirani, A.A.; Mirbahar, W.A.; Solangi, M.A.; Abul-Soad, A.A.; Markhand, G.S. Influence of Basal Salts, Sucrose and Plant Growth Regulator Levels on Nucellar Embryogenesis and Plantlet Regeneration in Monoembryonic Mango Varieties. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2023, 60, 431–441. [Google Scholar] [CrossRef]
- Lu, P.; Chen, J.; Tang, J.; Zhang, M. Research Progress on the Application of Plant Growth Regulators in Wheat Production. Barley Cereal Sci. 2023, 40, 1–7. [Google Scholar]
- Lad, B.L.; Jayasankar, S.; Pliego-Alfaro, F.; Moon, P.A.; Litz, R.E. Temporal effect of 2, 4-D on induction of embryogenic nuclellar cultures and somatic embryo development of ‘Carabao’mango. In Vitr. Cell. Dev. Biol.-Plant 1997, 33, 253–257. [Google Scholar] [CrossRef]
- Qureshi, M.A.; Shafqat, W.; ud Din, S.; Raza, M.K.; Sharif, N.; Ikram, S.; Rana, H.A.A. Effect of different plant growth regulators on in vitro regeneration of mango: A report. J. Glob. Innov. Agric. Sci. 2021, 9, 95–98. [Google Scholar]
- Huang, Y.f.; Zhou, G.L.; Zhao, H.F. Effect of different culture conditions on browning rate of explants in tissue culture of teatrees. J. Southwest. China J. Agric. Sci. 2009, 22, 1492–1495. [Google Scholar]
- Huang, L.; Wei, Z. Differences in regeneration ability and DNA of embryogenic and non embryogenic calli in different genotypes of Maize. Acta Phytophysiol. Sin. 1999, 25, 332–338. [Google Scholar]
- Ara, H.; Jaiswal, U.; Jaiswal, V.S. Plant regeneration from protoplasts of mango (Mangifera indica L.) through somatic embryogenesis. Plant Cell Rep. 2000, 19, 622–627. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Zhou, S.C.; Liu, Y.F.; Zhang, G.F. Influences of Plant Growth Regulations on Induction and Differentiation of Plant Callus. J. CUN (Nat. Sci. Ed.) 2007, 16, 23–28. [Google Scholar]
- Jiang, Y.; Liu, H.Z.; Li, K.L. Optimizing Populus ussuriensis Regeneration System. J. Northeast. For. Univ. 2017, 4, 28–32. [Google Scholar]
- Wu, J.Y.; Tian, X.C.; Xiao, S.Q.; Zhou, W.L.; Qiu, C.Y.; Li, W.D.; Liu, Y. Effects of Different Concentrations of Hormone on Stem Tissue Culture of Axillary Buds in Eucommia. Mol. Plant Breed. 2024, 1–10. [Google Scholar]
- Shekhawat, M.S.; Mehta, S.R.; Manokari, M.; Priyadharshini, S.; Badhepuri, M.K.; Jogam, P.; Dey, A.; Rajput, B.S. Morpho-anatomical and physiological changes of Indian sandalwood (Santalum album L.) plantlets in ex vitro conditions to support successful acclimatization for plant mass production. Plant Cell Tissue Organ. Cult. 2021, 147, 423–435. [Google Scholar] [CrossRef]
- Rivera-Domínguez, M.; Manzanilla-Ramírez, M.A.; Robles-González, M.; Gómez-Lim, M.A. Induction of somatic embryogenesis and plant regeneration of ‘Ataulfo’mango (Mangifera indica). Plant Cell Tissue Organ. Cult. 2004, 79, 101–104. [Google Scholar] [CrossRef]
- Litz, R.E.; Yurgalevitch, C. Effects of 1-aminocyclopropane-1-carboxylic acid, aminoethoxyvinylglycine, methylglyoxal bis-(guanylhydrazone) and dicyclohexylammonium sulfate on induction of embryogenic competence of mango nucellar explants. Plant Cell Tissue Organ. Cult. 1997, 51, 171–176. [Google Scholar] [CrossRef]
- Litz, R.E.; Hendrix, R.C.; Moon, P.A.; Chavez, V.M. Induction of embryogenic mango cultures as affected by genotype, explanting, 2,4-D and embryogenic nurse culture. Tissue Organ Cult. 1998, 53, 13–18. [Google Scholar] [CrossRef]
- .Monsalud, M.J.; Mathews, H.; Litz, R.E.; Gray, D.J. Control of hyperhydricity of mango somatic embryos. Plant Cell Tissue Organ. Cult. 1995, 42, 195–206. [Google Scholar] [CrossRef]
- Wang, X.F.; Fu, J.R.; Huang, H.L. Plant Regeneration from In Vitro Hypocotyls of Mango. J. Sun Yat Sen Univ. (Nat. Sci. Ed.) 1992, 31, 128–131. [Google Scholar]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propagators’ Soc. 1980, 30, 421–427. [Google Scholar]
- Falasca, G.; Zaghi, D.; Possenti, M.; Altamura, M.M. Adventitious root fomation in Arabidopsis thaliana thin cell layers. Plant Cell Rep. 2004, 23, 17–25. [Google Scholar] [CrossRef]
- Yu, J.; Liu, W.; Liu, J.; Qin, P.; Xu, L. Auxin Control of Root Organogenesis from Callus in Tissue Culture. Front. Plant Sci. 2017, 8, 1–4. [Google Scholar] [CrossRef]
- Xu, L. De novo root regeneration from leaf explants: Wounding, auxin, and cell fate transition. Curr. Opin. Plant Biol. 2018, 41, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Legué, V.; Rigal, A.; Bhalerao, R.P. Adventitious root formation in tree species: Involvement of transcription factors. Physiol. Plant. 2014, 151, 192–198. [Google Scholar] [CrossRef]
- Pateña, L.F.; Carlos-Refuerzo, L.R.; Barba, R.C. Somatic embryogenesis and plantlet regeneration in mango (Mangifera indica L.). Vitr. Cell. Dev. Biol.-Plant 2002, 38, 173–177. [Google Scholar] [CrossRef]
- Kumar, S.; Mangal, M.; Dhawan, A.K. Assessment of genetic fidelity of micropropagated plants of Simmondsia chinensis (Link) Schneider using RAPD and ISSR markers. Acta Physiol. Plant 2011, 33, 2541–2545. [Google Scholar] [CrossRef]
- Hou, J.; Su, P.; Wang, D.; Chen, X.; Zhao, W.; Wu, L. Efficient plant regeneration from in vitro leaves and petioles via shoot organogenesis in Sapium sebiferum Roxb. Plant Cell Tiss. Organ. Cult. 2020, 142, 143–156. [Google Scholar] [CrossRef]
- Saeed, T.; Shahzad, A.; Sharma, S. Studies on single and double layered biocompatible encapsulation of somatic embryos in Albizia lebbeck and genetic homogeneity appraisal among synseed derived lines through ISSR markers. Plant Cell Tissue Organ. Cult. 2020, 140, 431–445. [Google Scholar] [CrossRef]
- Rohela, G.K.; Jogam, P.; Mir, M.Y.; Shabnam, A.A.; Shukla, P.; Abbagani, S.; Kamili, A.N. Indirect regeneration and genetic fidelity analysis of acclimated plantlets through SCoT and ISSR markers in Morus alba L.cv. Chinese white. Biotechnol. Rep. 2020, 25, e00417. [Google Scholar]
- Murashige, T.; Skoog, F.K. A revised medium for the growth and bioassay with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D.J. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus lambertiana). Plant Cell Rep. 1985, 4, 177–179. [Google Scholar] [CrossRef]
- Gawel, N.J.; Jarret, R.L. A modified CTAB DNA extraction procedure for Musa and Ipomoea. Plant Mol. Bio. Rep. 1991, 9, 262–266. [Google Scholar]
- Li, Z.Q.; Dang, Z.G.; Zhao, Z.C.; Huang, J.F.; Gao, A.P.; Chen, Y.Y. Genetic Diversity Analyze of Mango’s F1 Hybrids and Construction of Genetic Map. Mol. Plant Breed. 2016, 14, 953–958. [Google Scholar]
- Chen, M. Development of CEAP Molecular Markers and Their Application in the Analysis of Mango Germplasm Resources; Guangxi University: Nanning, China, 2022. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 6-BA (mg/L) | NAA (mg/L) | Adventitious Shoot Induction Rate (%) | Average Number of Adventitious Shoots per Explant |
|---|---|---|---|
| 0.5 | 0.05 | 39.39 ± 0.48 f | 2.11 ± 0.32 e |
| 0.5 | 0.1 | 45.12 ± 1.27 e | 3.19 ± 0.60 d |
| 1 | 0.1 | 73.63 ± 0.55 a | 6.76 ± 0.64 a |
| 1 | 0.2 | 67.30 ± 0.83 b | 5.01 ± 0.58 b |
| 2 | 0.2 | 62.07 ± 1.11 c | 4.73 ± 0.60 c |
| 2 | 0.4 | 50.85 ± 1.61 d | 1.20 ± 0.52 f |
| Basal Medium | Adventitious Shoot Induction Rate (%) | Number of Adventitious Shoots per Explant |
|---|---|---|
| MS | 53.72 ± 0.89 c | 3.56 ± 0.73 c |
| 1/2 MS | 56.70 ± 1.31 b | 4.83 ± 0.52 b |
| WPM | 63.87 ± 1.14 a | 5.21 ± 0.69 a |
| DCR | 47.92 ± 0.74 d | 2.15 ± 0.54 d |
| 6-BA (mg/L) | Shoot Elongation (%) | Average Length (cm) | Description |
|---|---|---|---|
| 0.1 | 75.00 ± 1.73 b | 4.51 ± 0.09 b | Healthy green leaves |
| 0.5 | 89.33 ± 2.51 a | 5.17 ± 0.16 a | Healthy green leaves |
| 1.0 | 66.00 ± 2.08 c | 4.26 ± 0.10 c | Leaf edge coking |
| IBA (mg/L) | Mean Number of Roots per Explant | Adventitious Root Induction Rate (%) |
|---|---|---|
| 1 | 1.33 ± 0.06 d | 33.03 ± 0.15 d |
| 2 | 4.23 ± 0.25 b | 49.23 ± 0.51 b |
| 3 | 6.43 ± 0.26 a | 66.13 ± 0.35 a |
| 4 | 2.60 ± 0.17 c | 38.63 ± 0.59 c |
| Primer Code | Primer Sequence (5′–3′) | Annealing Temperature (°C) | No. of Scorable Bands | Size Range of Bands (bp) |
|---|---|---|---|---|
| UBC808 | AGAGAGAGAGAGAGAGC | 50 | 4 | 250–2000 |
| UBC836 | AGAGAGAGAGAGAGAGYA | 48 | 9 | 250–2000 |
| UBC848 | CACACACACACACACARG | 50 | 4 | 300–2000 |
| UBC855 | ACACACACACACACACYT | 48 | 7 | 300–1200 |
| UBC856 | ACACACACACACACACYA | 48 | 5 | 300–1800 |
| UBC886 | VDVCTCTCTCTCTCTCT | 45 | 8 | 250–2000 |
| UBC888 | BDBCACACACACACACA | 45 | 6 | 300–1500 |
| UBC890 | VHVGTGTGTGTGTGTGT | 45 | 5 | 300–1200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Sun, J.; Zheng, K.; Zhang, X.; Yao, Y.; Zhu, M. Efficient Plantlet Regeneration from Branches in Mangifera indica L. Plants 2024, 13, 2595. https://doi.org/10.3390/plants13182595
Zhou H, Sun J, Zheng K, Zhang X, Yao Y, Zhu M. Efficient Plantlet Regeneration from Branches in Mangifera indica L. Plants. 2024; 13(18):2595. https://doi.org/10.3390/plants13182595
Chicago/Turabian StyleZhou, Huijing, Jinglang Sun, Keyuan Zheng, Xinyuan Zhang, Yuan Yao, and Mulan Zhu. 2024. "Efficient Plantlet Regeneration from Branches in Mangifera indica L." Plants 13, no. 18: 2595. https://doi.org/10.3390/plants13182595
APA StyleZhou, H., Sun, J., Zheng, K., Zhang, X., Yao, Y., & Zhu, M. (2024). Efficient Plantlet Regeneration from Branches in Mangifera indica L. Plants, 13(18), 2595. https://doi.org/10.3390/plants13182595