
The Structural and Functional Responses of Rhizosphere Bacteria to Biodegradable Microplastics in the Presence of Biofertilizers



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
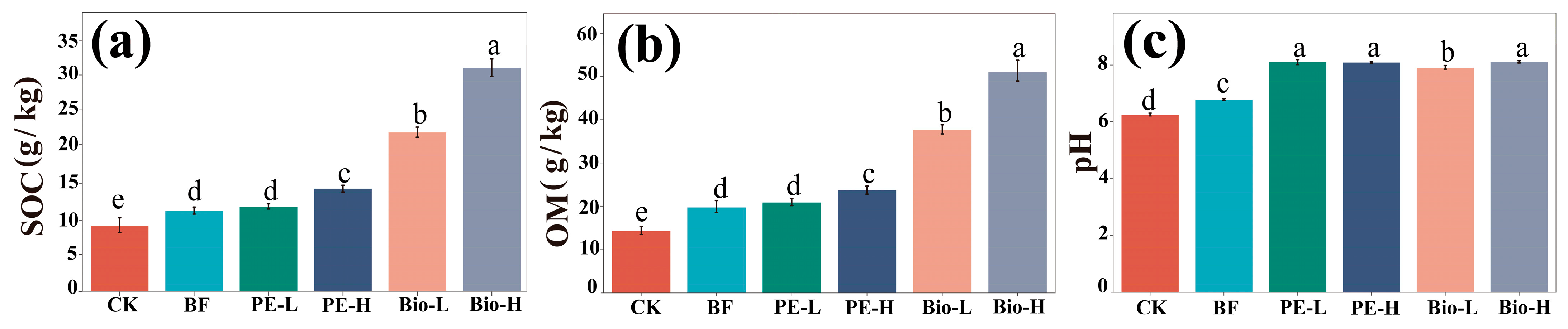
2.1. Changes in the Physicochemical Properties of the Rhizosphere
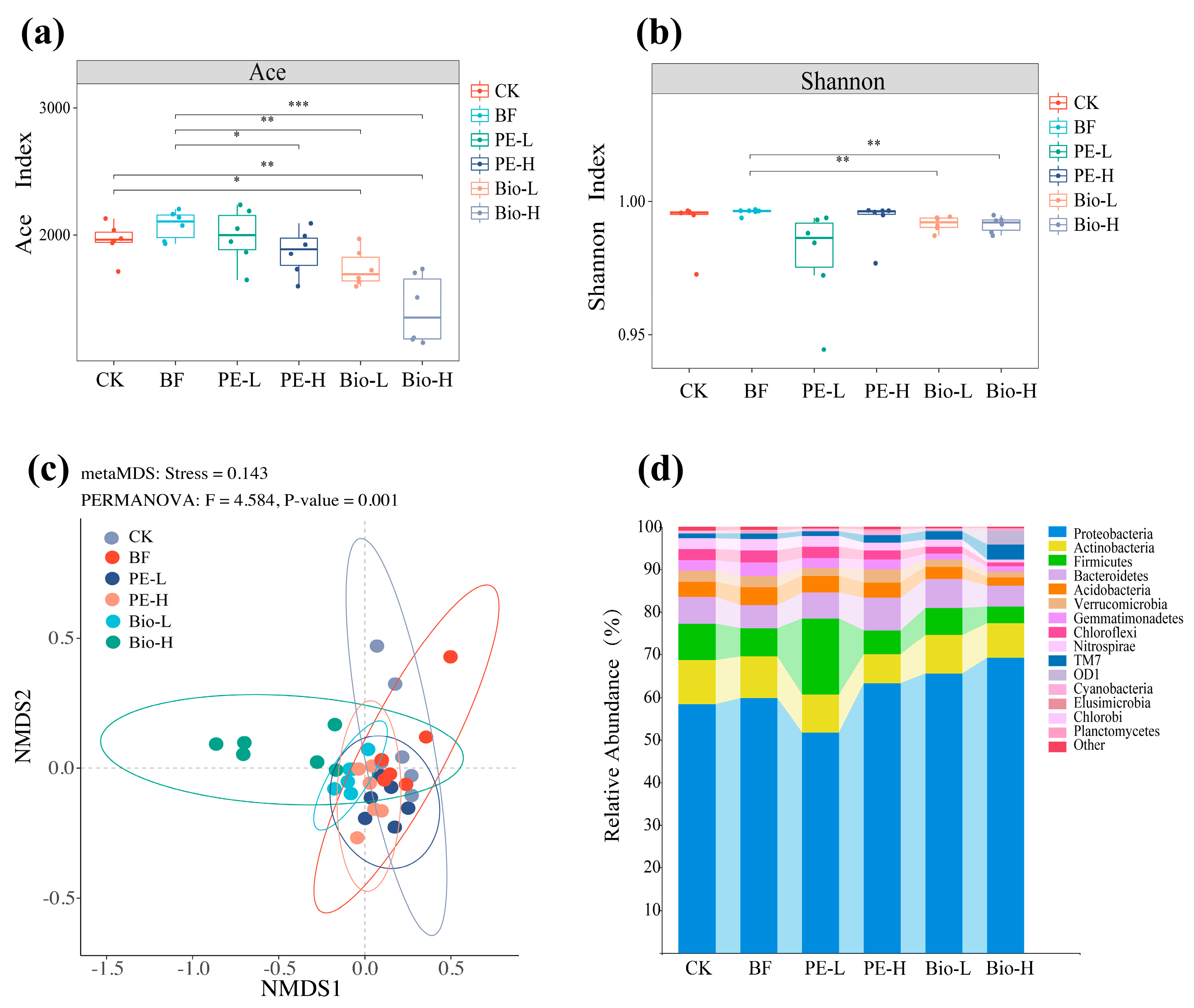
2.2. Changes in Bacterial Communities in the Rhizospheric Environment
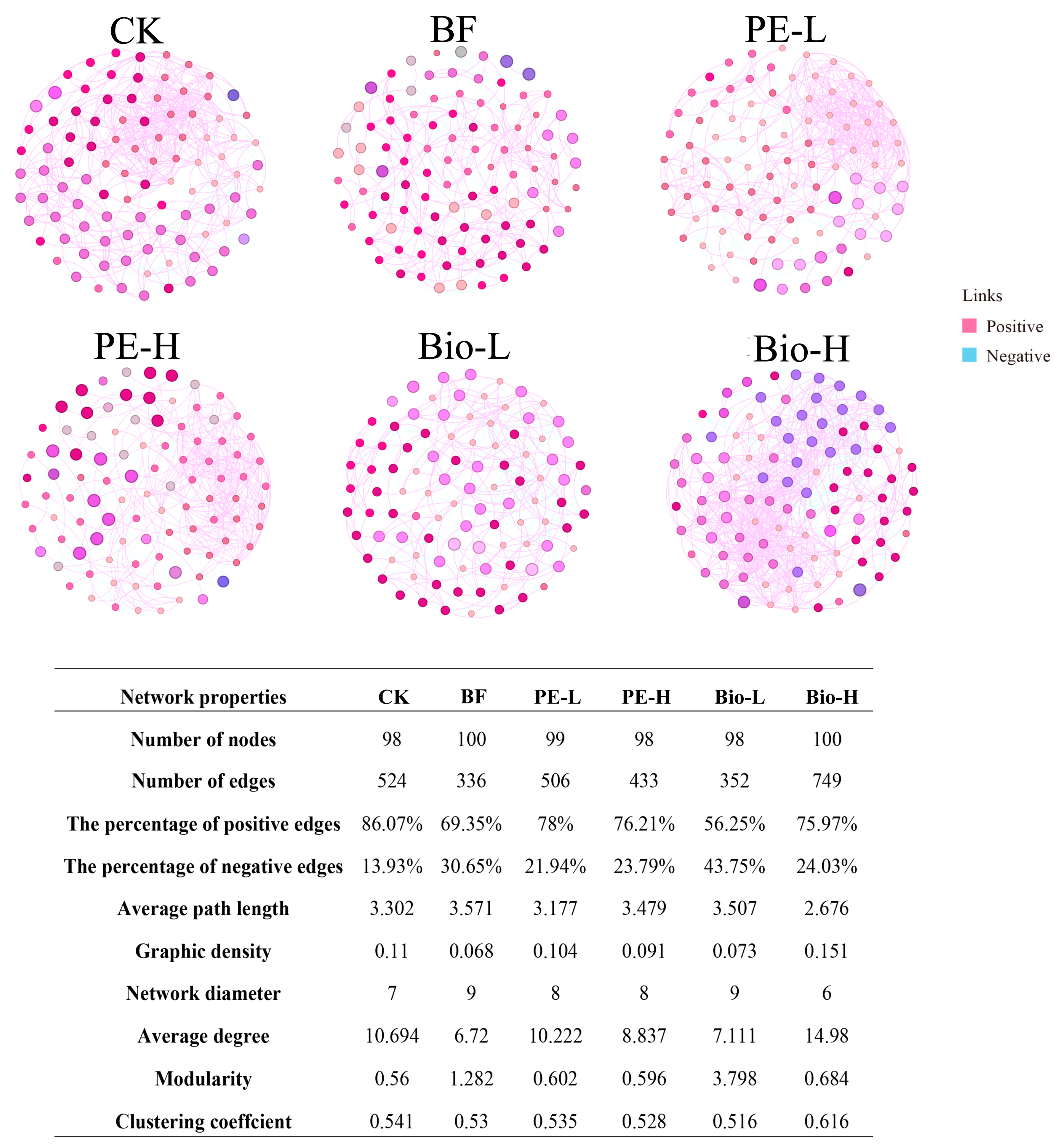
2.3. The Network Profiles of the Rhizosphere Bacterial Community
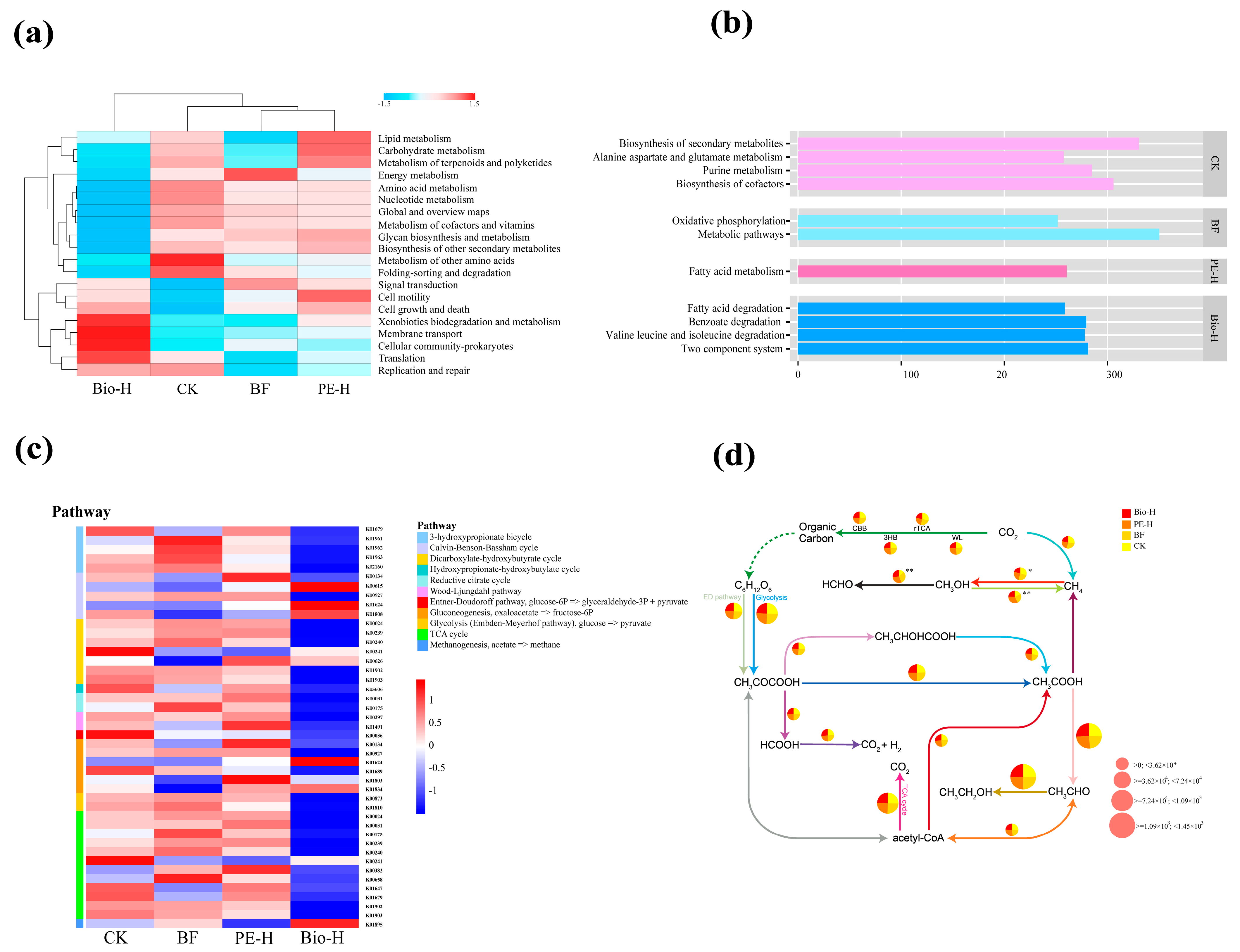
2.4. Rhizosphere Bacterial Function Based on Metagenomic Analysis
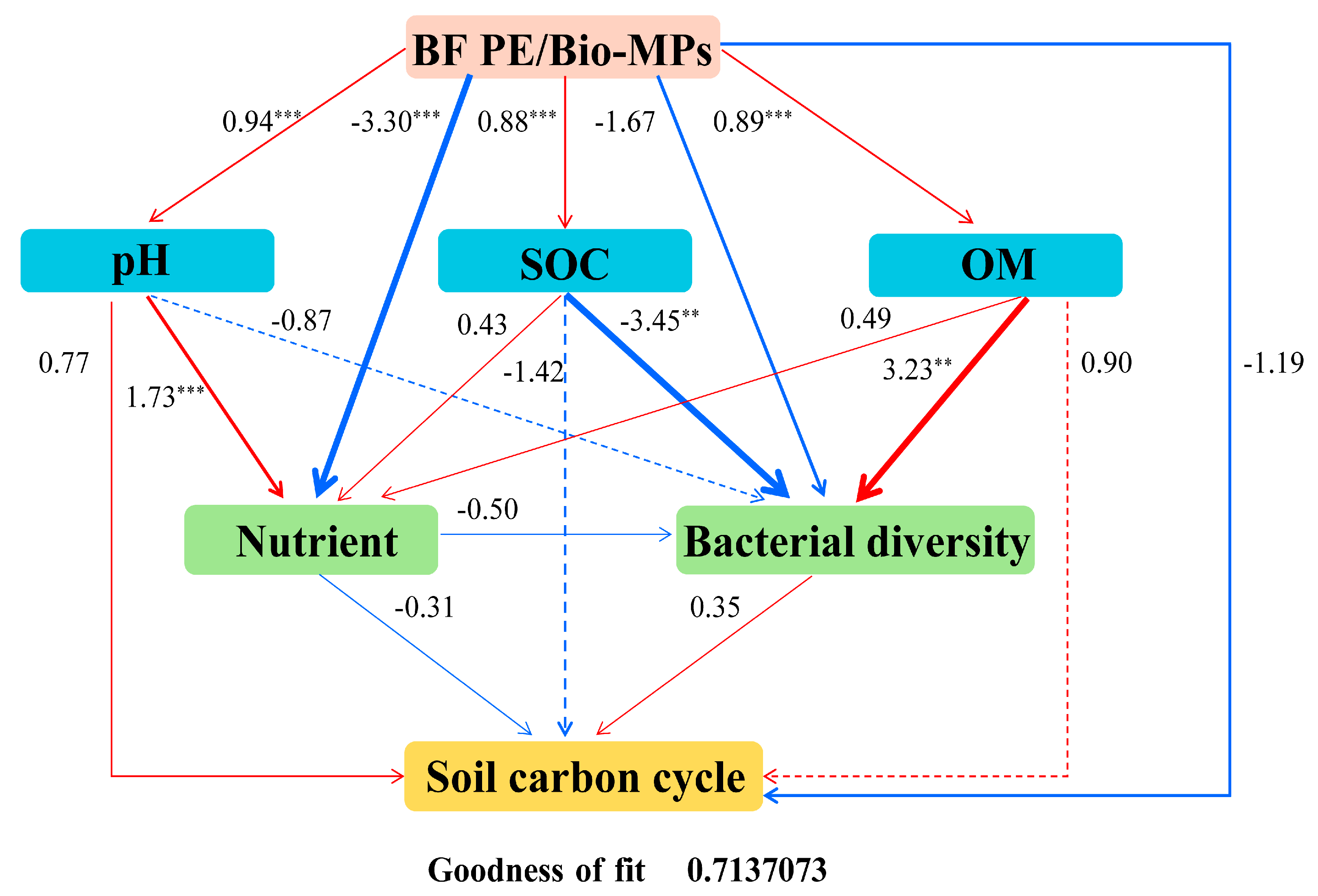
2.5. Partial Least Squares Path Model (PLS-PM) Analysis
3. Materials and Methods
3.1. Preparation of Biofertilizers and Characterization of MPs
3.2. Exposure Experiment of MPs
3.3. Physicochemical Properties of Rhizosphere Soil
3.4. DNA Extraction, PCR Amplification, and Metagenomic Analysis
3.5. Statistical Analyses
4. Discussion
4.1. Bacterial Composition Responses to Biofertilizers and MPs
4.2. Rhizosphere Bacterial Function Responses to Biofertilizers and MPs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility. plant tolerance and crop productivity. Microb. Cell Fact. 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2016, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Nosheen, S.; Ajmal, I.; Song, Y.D. Microbes as biofertilizers, a potential approach for sustainable crop production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Singh, J.S.; Pandey, V.C.; Singh, D.P. Efficient soil microorganisms: A new dimension for sustainable agriculture and environmental development. Agric. Ecosyst. Environ. 2011, 140, 339–353. [Google Scholar] [CrossRef]
- Ibáñez, A.; Diez-Galán, A.; Cobos, R.; Calvo-Peña, C.; Barreiro, C.; Medina-Turienzo, J.; Sánchez-García, M.; Coque, J.J.R. Using Rhizosphere Phosphate Solubilizing Bacteria to Improve Barley (Hordeum vulgare) Plant Productivity. Microorganisms. 2021, 9, 1619. [Google Scholar] [CrossRef]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of microplastics in soil ecosystems: Above and below ground. Environ. Sci. Technol. 2019, 53, 11496–11506. [Google Scholar] [CrossRef]
- Fuller, S.; Gautam, A. A procedure for measuring microplastics using pressurized fluid extraction. Environ. Sci. Technol. 2016, 50, 5774–5780. [Google Scholar] [CrossRef]
- Luca, N.; Martyn, F.; Sindre, L. Are agricultural soils dumps for microplastics of urban origin? Environ. Sci. Technol. 2016, 50, 10777–10779. [Google Scholar]
- Montazer, Z.; Habibi Najafi, M.B.; Levin, D.B. Challenges with verifying microbial degradation of polyethylene. Polymers 2020, 12, 123. [Google Scholar] [CrossRef]
- Silvia, C.; Riccardo, F.; Majid, G.; Jonan, M.A.; Stefano, L. Exploring sustainable alternatives: Wood distillate alleviates the impact of bioplastic in basil plants. Sci. Total Environ. 2023, 900, 166484. [Google Scholar]
- Azarnejad, N.; Celletti, S.; Ghorbani, M.; Fedeli, R.; Loppi, S. Dose-Dependent Effects of a Corn Starch-Based Bioplastic on Basil (Ocimum basilicum L.): Implications for Growth, Biochemical Parameters, and Nutrient Content. Toxics 2024, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Yu, H.; Xi, B.D.; Tan, W.B. A review on the occurrence and influence of biodegradable microplastics in soil ecosystems: Are biodegradable plastics substitute or threat? Environ. Int. 2022, 163, 107244. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Chen, C.Y.; Song, B.; Shen, M.C.; Cao, W.C.; Yang, H.L.; Zeng, G.M.; Gong, J.L. A review of biodegradable plastics to biodegradable microplastics: Another ecological threat to soil environments? J. Cleaner Prod. 2021, 312, 127816. [Google Scholar] [CrossRef]
- Rodrigo, M.; Paolina, G.; Jos, M. Raaijmakers The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar]
- Dilfuza, E.; Faina, K.; Shamil, V.; Laziza, G.; Zulfiya, K.; Ben, L. High incidence of plant growth-stimulating bacteria associated with the rhizosphere of wheat grown on salinated soil in Uzbekistan. Environ. Microbiol. 2007, 10, 1–9. [Google Scholar]
- Amanda, A.G.; Paula, M.D.J.; Rafaela, S.A.N.; Ligiane, A.F.; Karina, B.S.; Fatima, M.S.M. Genetic and symbiotic diversity of nitrogen-fixing bacteria isolated from agricultural soils in the western amazon by using cowpea as the trap plant. Appl. Environ. Microbiol. 2012, 78, 6726–6733. [Google Scholar]
- Petra, M.; David, C.; Zed, R. Rhizosphere interactions between microorganisms and plants govern iron and phosphorus acquisition along the root axis-model and research methods. Soil Biol. Biochem. 2011, 43, 883–894. [Google Scholar]
- Collignon, C.; Uroz, S.; Turpault, M.P.; Frey-Klett, P. Seasons differently impact the structure of mineral weathering bacterial communities in beech and spruce stands. Soil Biol. Biochem. 2011, 43, 2012–2022. [Google Scholar] [CrossRef]
- Christos, Z.; Corné, M.J.P. Modulation of host immunity by beneficial microbes. Mol. Plant-Microbe Interact. 2012, 25, 139–150. [Google Scholar]
- Li, C.D.; Cui, Q.; Li, Y.; Zhang, K.; Lu, X.; Zhang, Y. Effect of LDPE and biodegradable PBAT primary microplastics on bacterial community after four months of soil incubation. J. Hazard. Mater. 2022, 429, 128353. [Google Scholar] [CrossRef]
- Michaela, K.R.; Lucy, M.G.; Martine, G.; Charlotte, E.M.L.; Richard, P.E.; Dave, R.C.; Davey, L.J. LDPE and biodegradable PLA-PBAT plastics differentially affect plant-soil nitrogen partitioning and dynamics in a Hordeum vulgare mesocosm. J. Hazard. Mater. 2023, 447, 130825. [Google Scholar]
- Wang, T.; Cheng, K.K.; Huo, X.J.; Meng, P.P.; Cai, Z.H.; Wang, Z.K.; Zhou, J. Bioorganic fertilizer promotes pakchoi growth and shapes the soil microbial structure. Front. Plant Sci. 2022, 13, 1040437. [Google Scholar] [CrossRef]
- Sreejata, B.; Lluis, M.C.; Ana, M.P.; Jennifer, M.D. Biodegradable plastic mulch films: Impacts on soil microbial communities and ecosystem functions. Front. Microbiol. 2018, 9, 819. [Google Scholar]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and soil microbial community: A review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- De Souza Machado, A.A.; Lau, C.W.; Till, J.; Kloas, W.; Lehmann, A.; Becker, R.; Rillig, M.C. Impacts of microplastics on the soil biophysical environment. Environ. Sci. Technol. 2018, 52, 9656–9665. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.F.; Yang, X.M.; Liu, G.B.; Liang, C.T.; Xue, S.; Chen, H.; Ritsema, C.J.; Geissen, V. Response of soil dissolved organic matter to microplastic addition in Chinese loess soil. Chemosphere 2017, 185, 907–917. [Google Scholar] [CrossRef]
- Xiao, H.; Liu, G.; Liu, P.L.; Zheng, F.L.; Zhang, J.Q.; Hu, F.N. Sediment transport capacity of concentrated flows on steep loessial slope with erodible beds. Sci. Rep. 2017, 7, 2350. [Google Scholar] [CrossRef]
- He, K.; Li, X.P.; Dong, L.L. The effects of flue gas desulfurization gypsum (FGD gypsum) on P fractions in a coastal plain soil. J. Soils Sediments. 2017, 18, 804–815. [Google Scholar] [CrossRef]
- Jiang, W.T.; Wang, X.K.; Xing, Y.Y.; Liu, X.H.; Cui, Z.G.; Yang, L.H. Enhancing rice production by potassium management: Recommended reasonable fertilization strategies in different inherent soil productivity levels for a sustainable rice production system. Sustainability 2019, 11, 6522. [Google Scholar] [CrossRef]
- Zhou, J.; Jia, R.; Brown, R.W.; Yang, Y.D.; Zeng, Z.H.; Jones, D.L.; Zang, H.D. The long-term uncertainty of biodegradable mulch film residues and associated microplastics pollution on plant-soil health. J. Hazard. Mater. 2023, 44, 130055. [Google Scholar] [CrossRef]
- Liu, D.; Iqbal, S.; Gui, H.; Xu, J.C.; An, S.; Xing, B.S. Nano-iron oxide (Fe3O4) mitigates the effects of microplastics on a ryegrass soil-microbe-plant system. ACS Nano. 2024, 17, 24867–24882. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Ma, J.; Li, G.; Rillig, M.C.; Zhu, Y.G. Soil plastispheres as hotpots of antibiotic resistance genes and potential pathogens. ISME J. 2021, 16, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Zhao, Y.R.; Qin, X.; Jia, W.Q.; Chai, L.W.; Huang, M.K.; Huang, Y. Microplastics from mulching film is a distinct habitat for bacteria in farmland soil. Sci. Total Environ. 2019, 688, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Ghosh, E.; Mukherjee, P.; Gajendiran, A. Microbial degradation of low density polyethylene. Environ. Prog. Sustainable Energy 2016, 36, 147–154. [Google Scholar] [CrossRef]
- Muhonja, C.N.; Magoma, G.; Imbuga, M.; Makonde, H.M. Molecular characterization of low-density polyethene (LDPE) degrading bacteria and fungi from Dandora Dumpsite, Nairobi, Kenya. Int. J. Microbiol. 2018, 2018, 4167845. [Google Scholar]
- Fei, Y.F.; Huang, S.Y.; Zhang, H.B.; Tong, Y.Z.; Wen, D.S.; Xia, X.Y.; Wang, H.; Luo, Y.M.; Barceló, D. Response of soil enzyme activities and bacterial communities to the accumulation of microplastics in an acid cropped soil. Sci. Total Environ. 2019, 707, 135634. [Google Scholar] [CrossRef]
- Ali, N.; Khan, S.; Li, Y.Y.; Zheng, N.G.; Yao, H.Y. Influence of biochars on the accessibility of organochlorine pesticides and microbial community in contaminated soils. Sci. Total Environ. 2019, 647, 551–560. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Chen, H.Y.; Wang, Q.; Liu, T.; Duan, Y.M.; Awasthi, S.K.; Ren, X.N.; Tu, Z.N.; Li, J.; Zhao, J.; et al. Succession of bacteria diversity in the poultry manure composted mixed with clay: Studies upon its dynamics and associations with physicochemical and gaseous parameters. Bioresour. Technol. 2018, 267, 618–625. [Google Scholar] [CrossRef]
- Liang, J.; Tang, S.Q.; Gong, J.L.; Zeng, G.M.; Tang, W.W.; Song, B.; Zhang, P.; Yang, Z.X.; Luo, Y. Responses of enzymatic activity and microbial communities to biochar/compost amendment in sulfamethoxazole polluted wetland soil. J. Hazard. Mater. 2019, 385, 121533. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.C.; Wang, H.Y.; Gai, X.P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Elad, Y.; Paudel, I.; Graber, E.R.; Cytryn, E.; Frenkel, O. Linking the Belowground Microbial Composition, Diversity and Activity to Soilborne Disease Suppression and Growth Promotion of Tomato Amended with Biochar. Sci. Rep. 2017, 7, 44382. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Qu, Y.Y.; Shen, W.L.; Zhang, Z.J.; Wang, J.W.; Liu, Z.Y.; Li, D.X.; Li, H.J.; Zhou, J.T. Bacterial community compositions of coking wastewater treatment plants in steel industry revealed by Illumina high-throughput sequencing. Bioresour. Technol. 2015, 179, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Dammak, M.; Fourati, Y.; Tarrés, Q.; Delgado-Aguilar, M.; Mutjé, P.; Boufi, S. Blends of PBAT with plasticized starch for packaging applications: Mechanical properties, rheological behaviour and biodegradability. Ind. Crops Prod. 2020, 144, 112061. [Google Scholar] [CrossRef]
- She, J.Y.; Liu, J.; He, H.P.; Zhang, Q.; Lin, Y.Y.; Wang, J.; Yin, M.L.; Wang, L.L.; Wei, X.D.; Huang, Y.L.; et al. Microbial response and adaption to thallium contamination in soil profiles. J. Hazard. Mater. 2020, 423, 127080. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Li, P.F.; Liu, J.; Saleem, M.; Li, G.L.; Luan, L.; Wu, M.; Li, Z.P. Reduced chemodiversity suppresses rhizosphere microbiome functioning in the mono-cropped agroecosystems. Microbiome. 2022, 10, 108. [Google Scholar] [CrossRef]
- Xiong, X.; Liao, H.; Xing, Y.F.; Han, X.K.; Wang, W.L.; Wan, W.J.; Huang, Q.Y.; Chen, W.L. Soil aggregates and fertilizer treatments drive bacterial interactions via interspecies niche overlap. Microbiol. Spectrum. 2022, 10, e02524-21. [Google Scholar] [CrossRef]
- Wu, C.C.; Ma, Y.J.; Wang, D.; Shan, Y.P.; Song, X.P.; Hu, H.Y.; Ren, X.L.; Ma, X.Y.; Cui, J.J.; Ma, Y. Integrated microbiology and metabolomics analysis reveal plastic mulch film residue affects soil microorganisms and their metabolic functions. J. Hazard. Mater. 2022, 423, 127258. [Google Scholar] [CrossRef]
- Ma, R.J.; Xu, Z.N.; Sun, J.Y.; Li, D.R.; Cheng, Z.; Niu, Y.L.; Guo, H.; Zhou, J.; Wang, T.C. Microplastics affect C, N, and P cycling in natural environments: Highlighting the driver of soil hydraulic properties. J. Hazard. Mater. 2023, 459, 132326. [Google Scholar] [CrossRef]
- Mai, Z.M.; Ye, M.; Wang, Y.S.; Foong, S.W.; Wang, L.; Sun, F.L.; Cheng, H. Characteristics of microbial community and function with the succession of mangroves. Front. Microbiol. 2021, 12, 764974. [Google Scholar] [CrossRef]
- Choi, G.M.; Im, W.T. Paraburkholderia azotifigens sp. nov., a nitrogen-fixing bacterium isolated from paddy soil. Int. J. Syst. Evol. Microbiol. 2018, 68, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, J.H.; Jia, L.; Gu, H.Y. Integrating metagenomics and metabolomics to study the response of microbiota in black soil degradation. Sci. Total Environ. 2023, 899, 165486. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.S.; Yan, R.; Liu, D.L.; Jiang, S.W.; Cui, L.; Guo, X.W.; Wang, X.J.; Zhang, J.; Xiang, W.S. Trinickia diaoshuihuensis sp. nov., a plant growth promoting bacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Benitez, M.S.; Gardener, B.B.M. Linking Sequence to Function in Soil Bacteria: Sequence-Directed Isolation of Novel Bacteria Contributing to Soilborne Plant Disease Suppression. Appl. Environ. Microbiol. 2008, 75, 915–924. [Google Scholar] [CrossRef]
- Rodgers-Vieira, E.A.; Zhang, Z.F.; Adrion, A.C.; Gold, A.; Aitken, M.D. Identification of Anthraquinone-Degrading Bacteria in Soil Contaminated with Polycyclic Aromatic Hydrocarbons. Appl. Environ. Microbiol. 2015, 81, 3775–3781. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Li, X.; Cai, Z.; Wang, Z.; Zhou, J. The Structural and Functional Responses of Rhizosphere Bacteria to Biodegradable Microplastics in the Presence of Biofertilizers. Plants 2024, 13, 2627. https://doi.org/10.3390/plants13182627
Cheng X, Li X, Cai Z, Wang Z, Zhou J. The Structural and Functional Responses of Rhizosphere Bacteria to Biodegradable Microplastics in the Presence of Biofertilizers. Plants. 2024; 13(18):2627. https://doi.org/10.3390/plants13182627
Chicago/Turabian StyleCheng, Xueyu, Xinyang Li, Zhonghua Cai, Zongkang Wang, and Jin Zhou. 2024. "The Structural and Functional Responses of Rhizosphere Bacteria to Biodegradable Microplastics in the Presence of Biofertilizers" Plants 13, no. 18: 2627. https://doi.org/10.3390/plants13182627