
Transcriptome Analysis for Salt-Responsive Genes in Two Different Alfalfa (Medicago sativa L.) Cultivars and Functional Analysis of MsHPCA1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
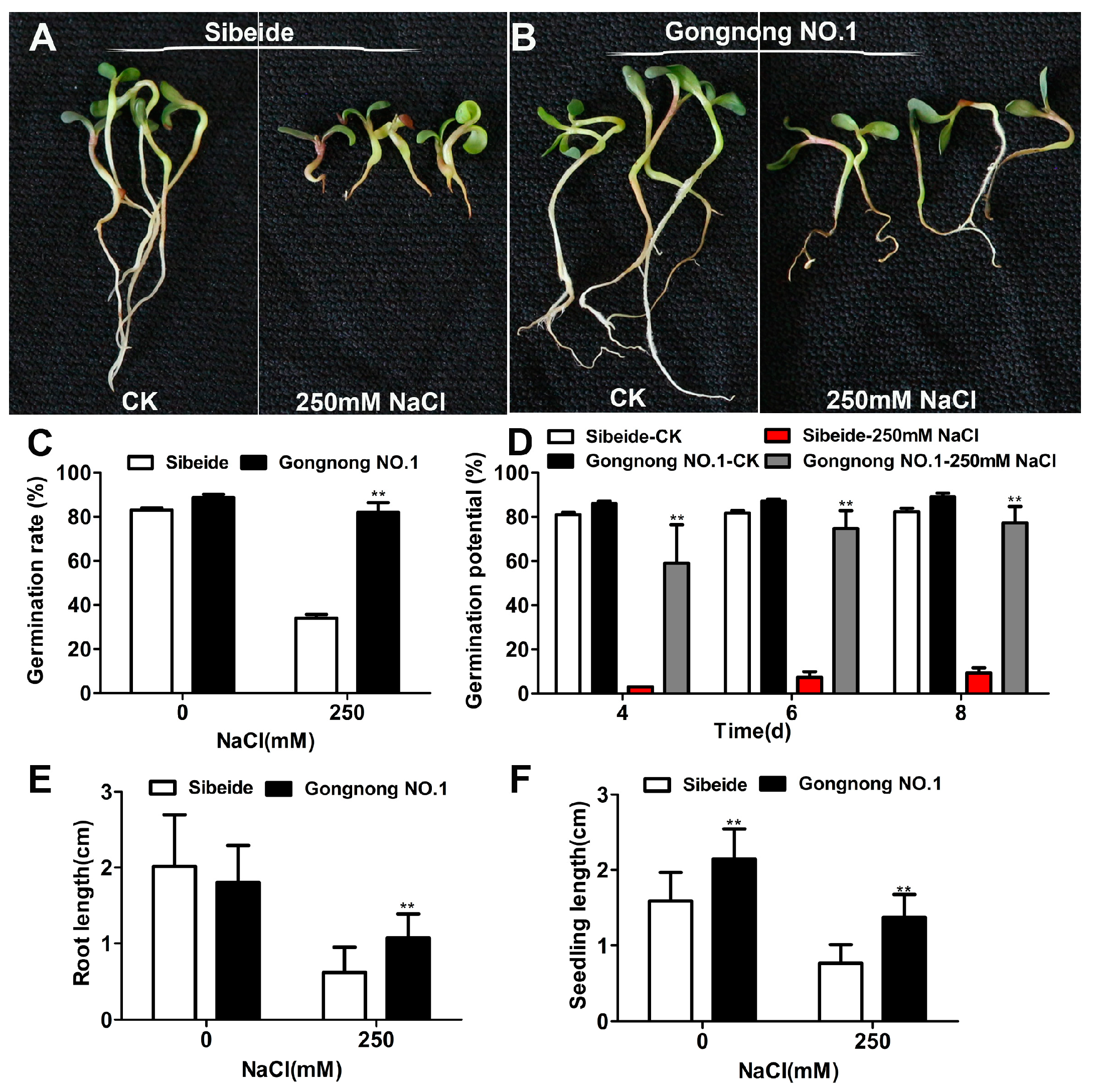
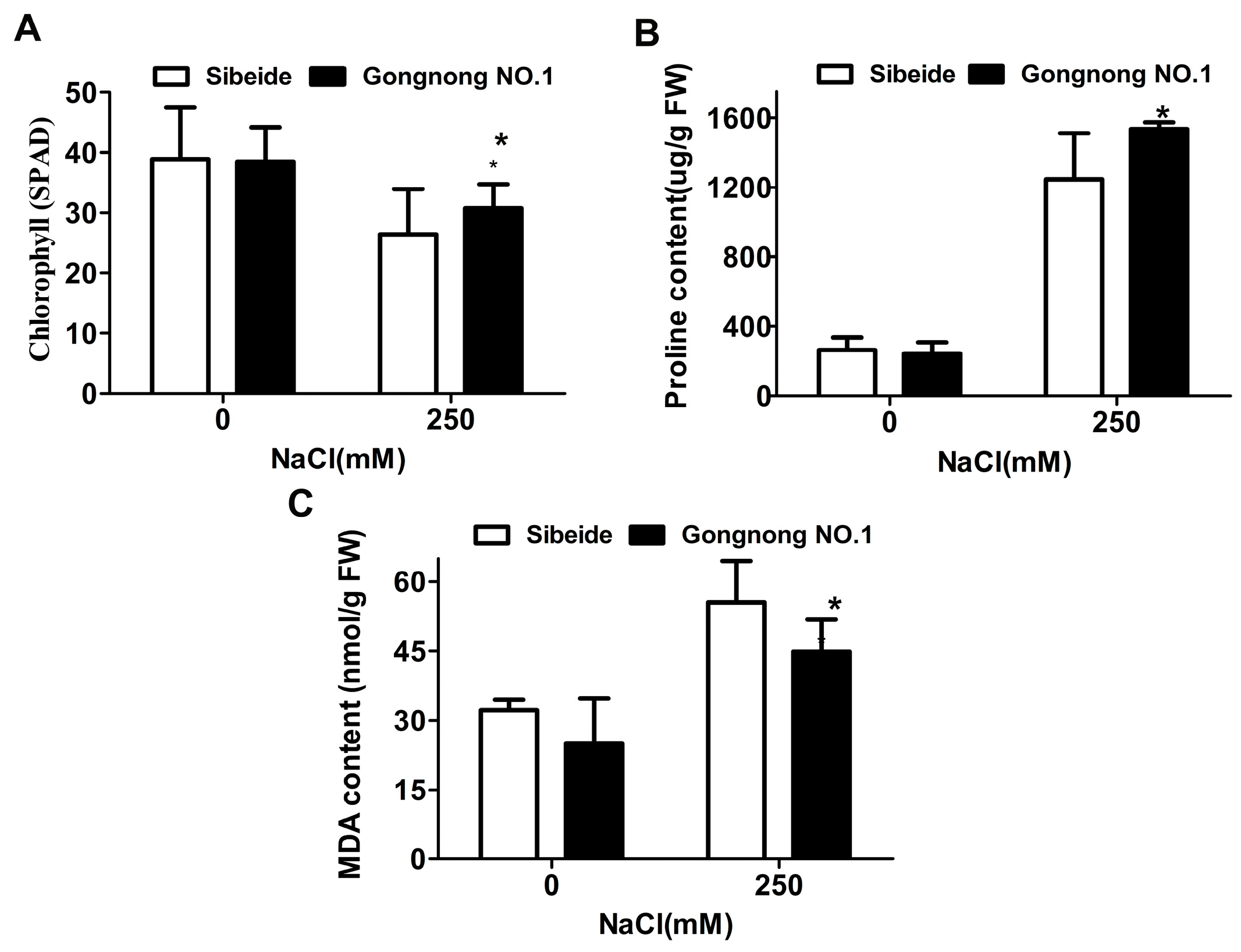
2.1. Physiological Responses of Alfalfa (‘Gongnong NO.1’ and ‘Sibeide’) toward NaCl Treatment
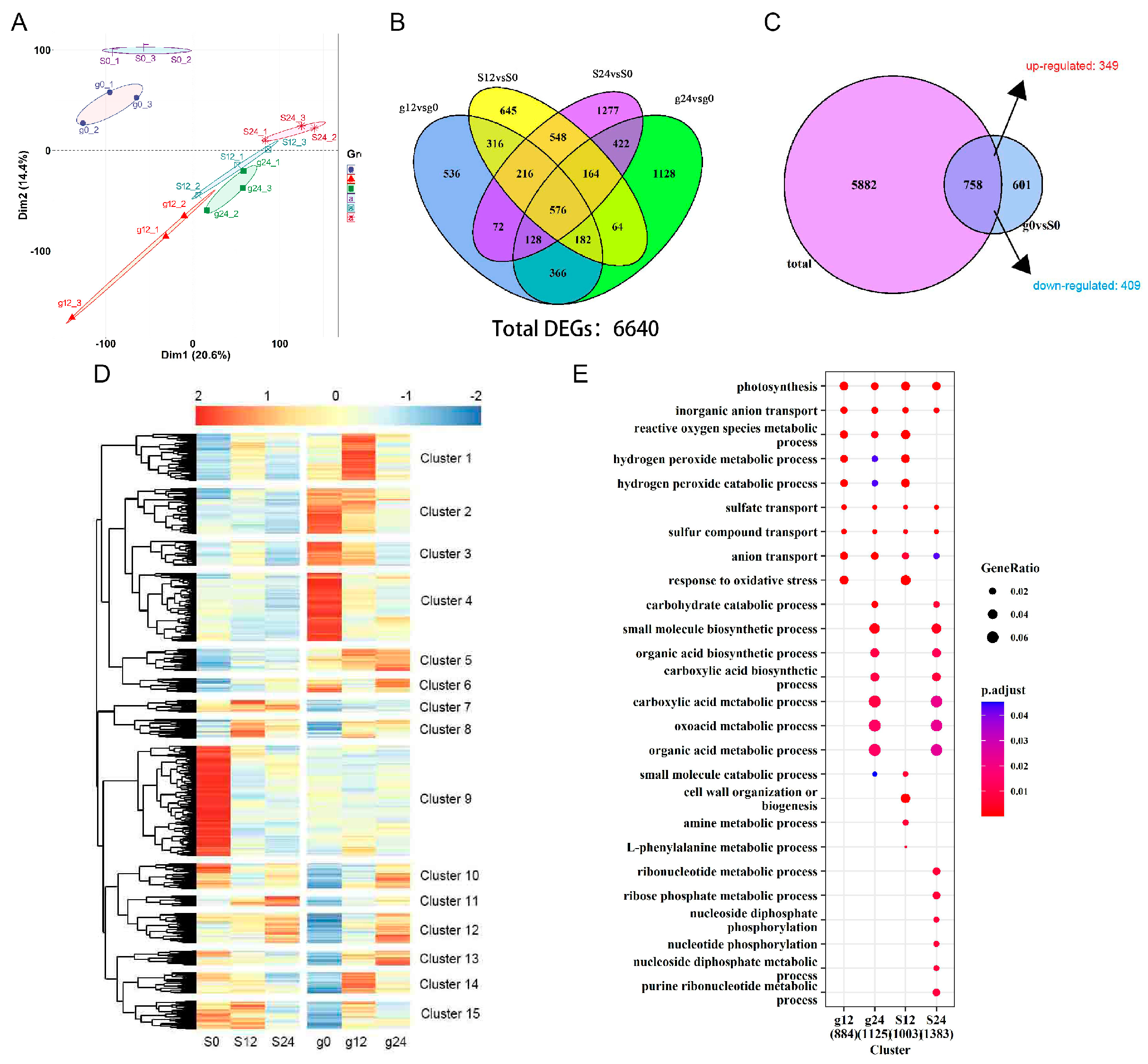
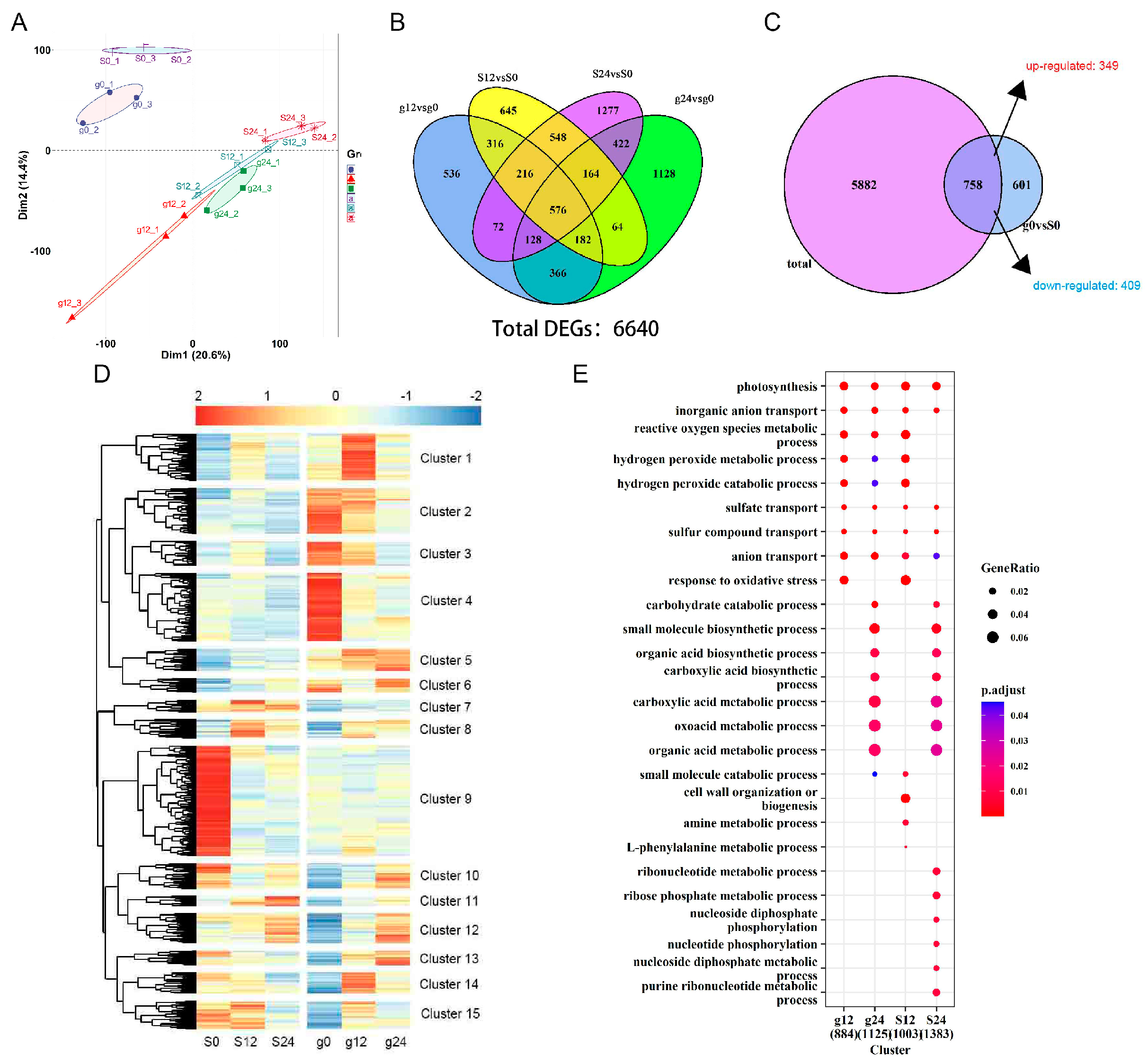
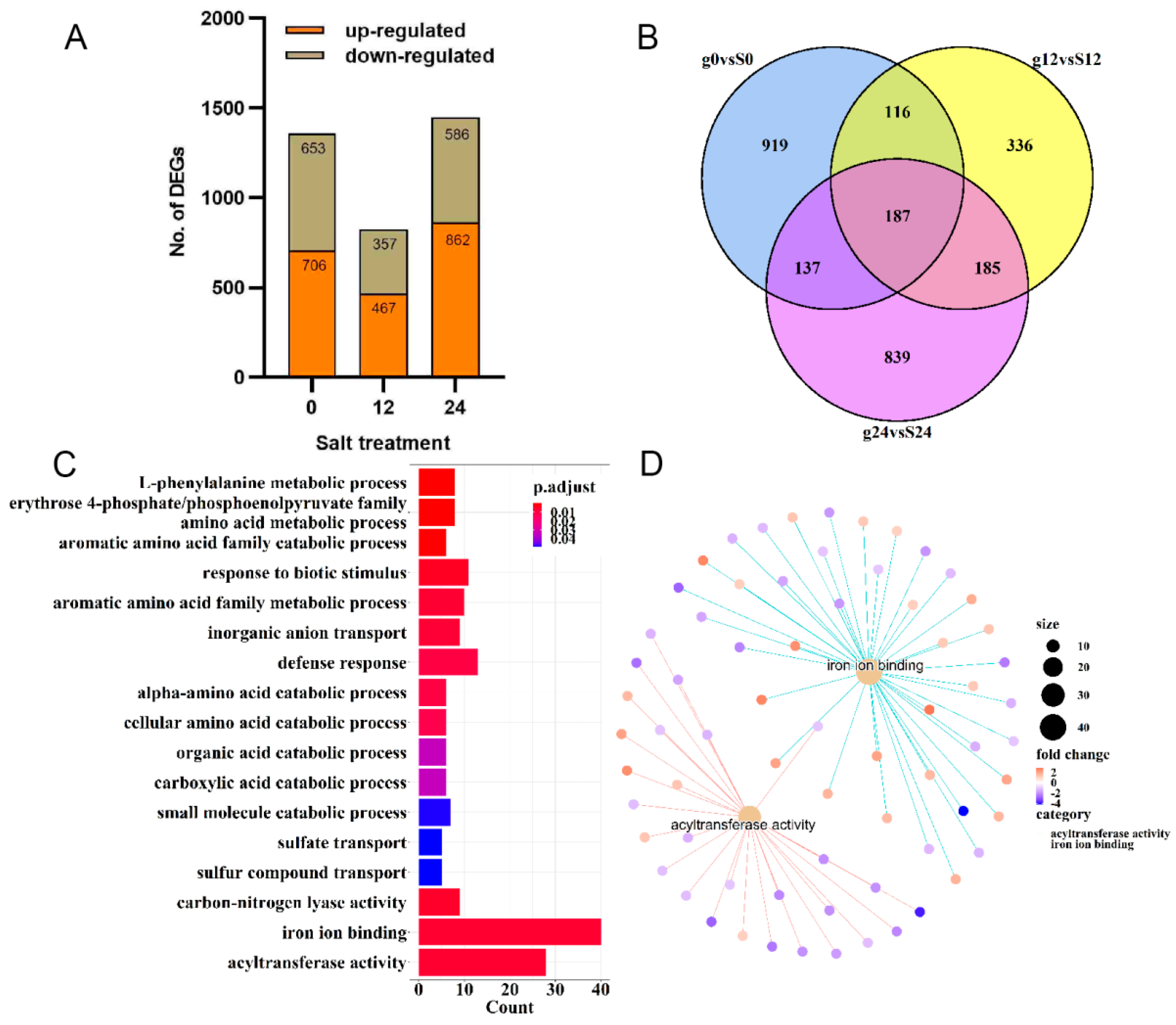
2.2. Identification of Differentially Expressed Genes in ‘Gongnong NO.1’ and ‘Sibeide’
2.3. Transcriptome Analysis of Differentially Expressed Genes in ‘Gongnong NO.1’ and ‘Sibeide’ under Salt Treatment
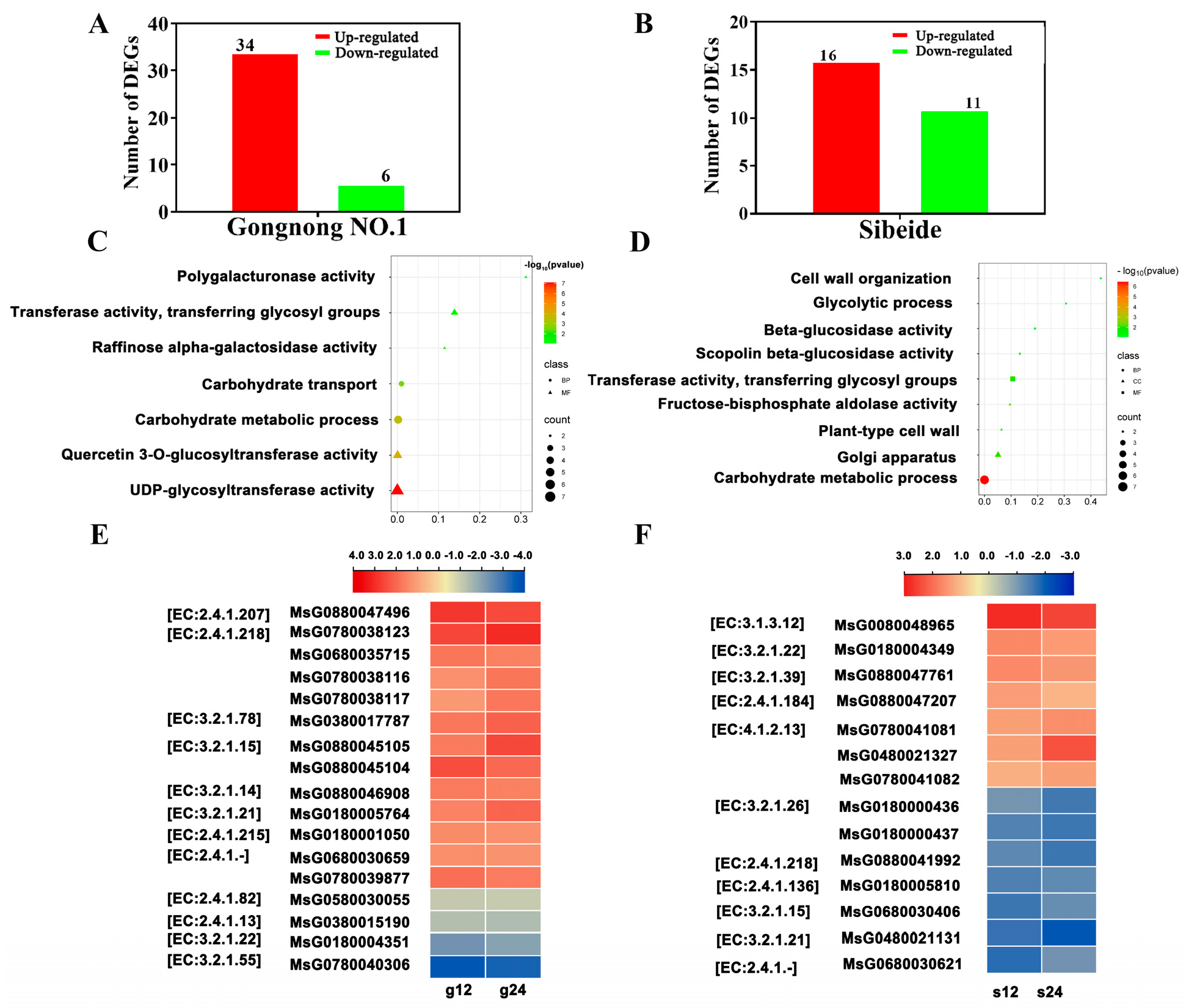
2.4. Analysis of Differentially Expressed Genes under Salt Treatment at Different Times
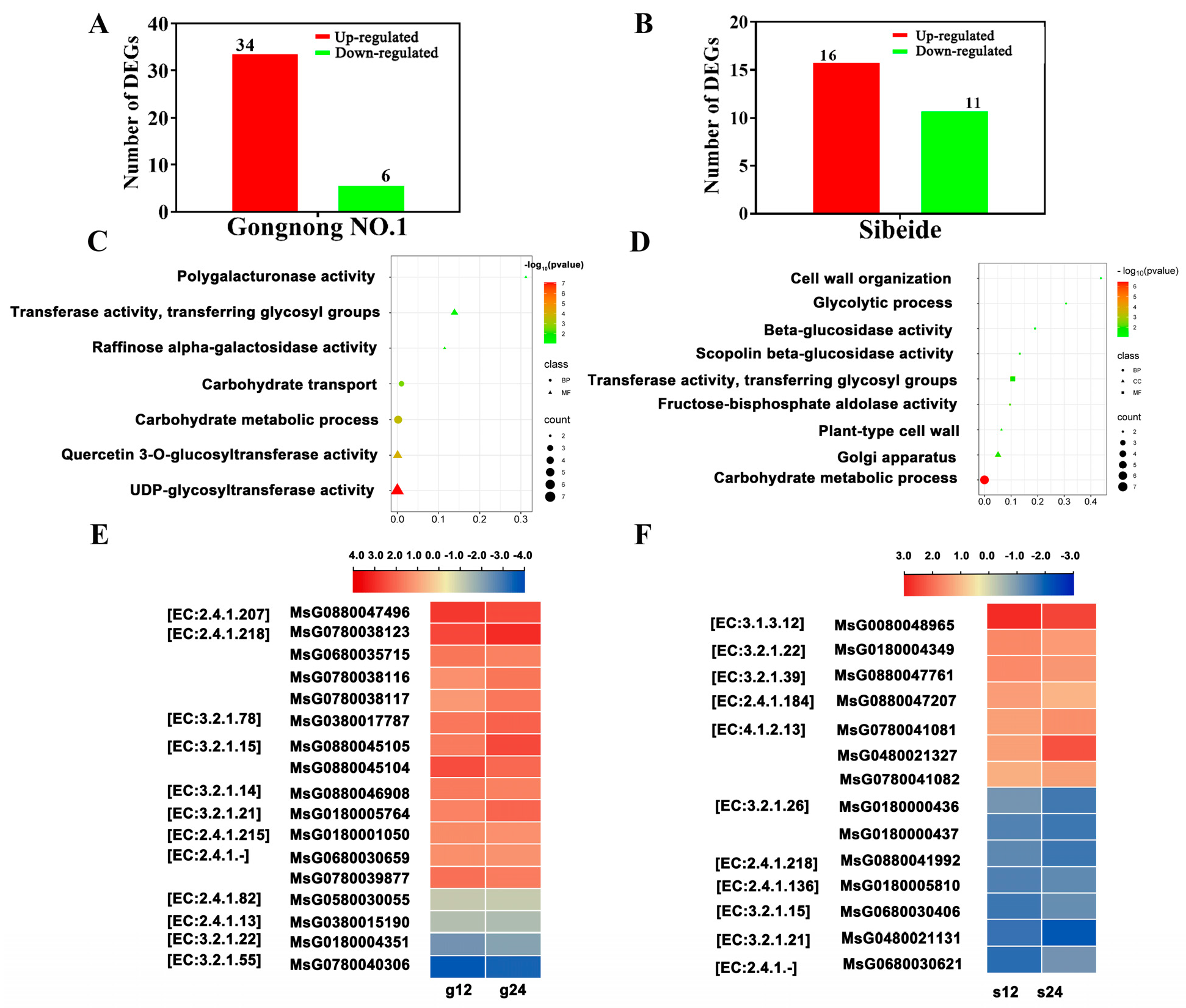
2.4.1. Glyco-Metabolism-Associated Gene
2.4.2. Energy-Associated Genes
2.4.3. Transcription Factor
2.4.4. Stress-Associated Gene
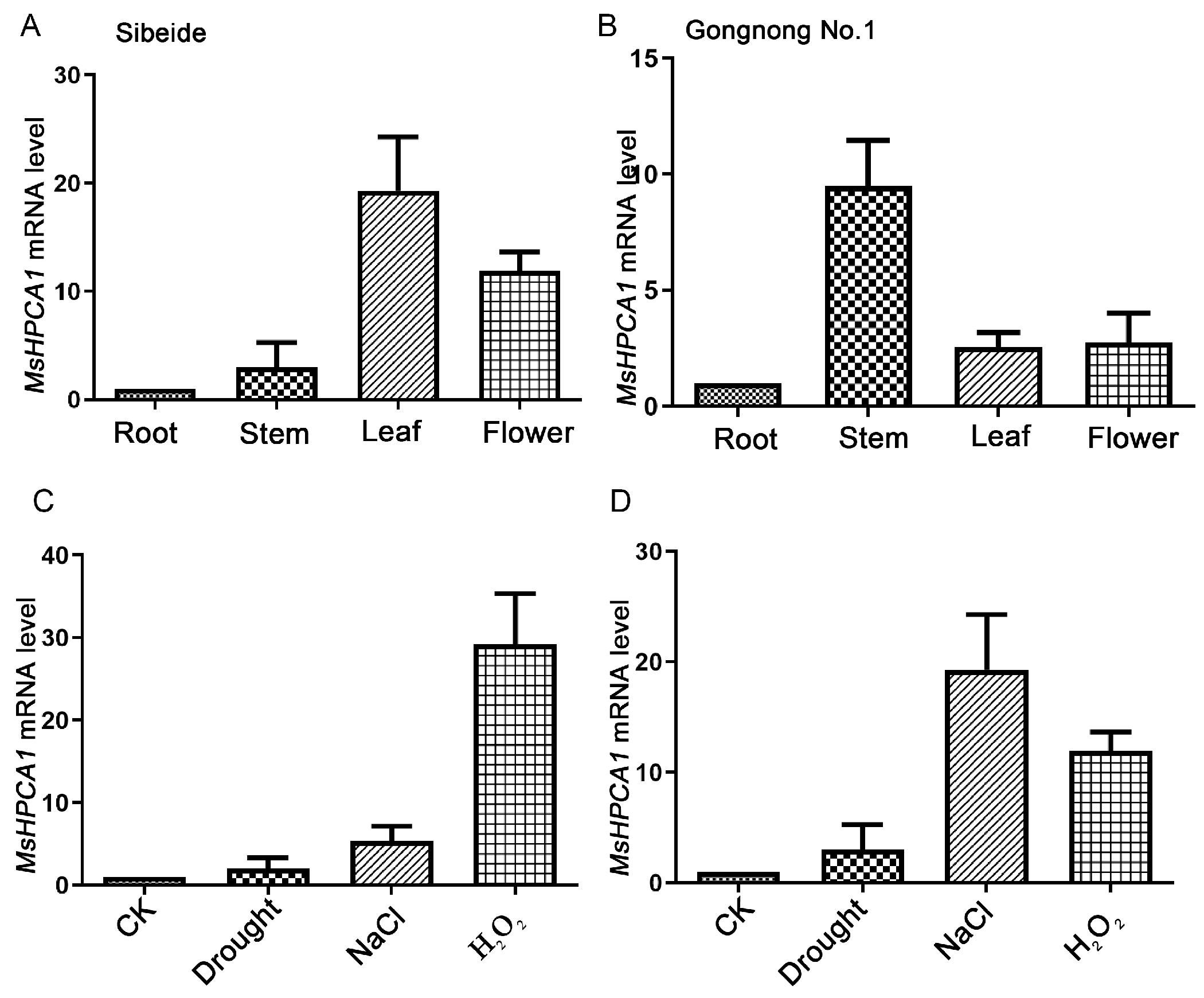
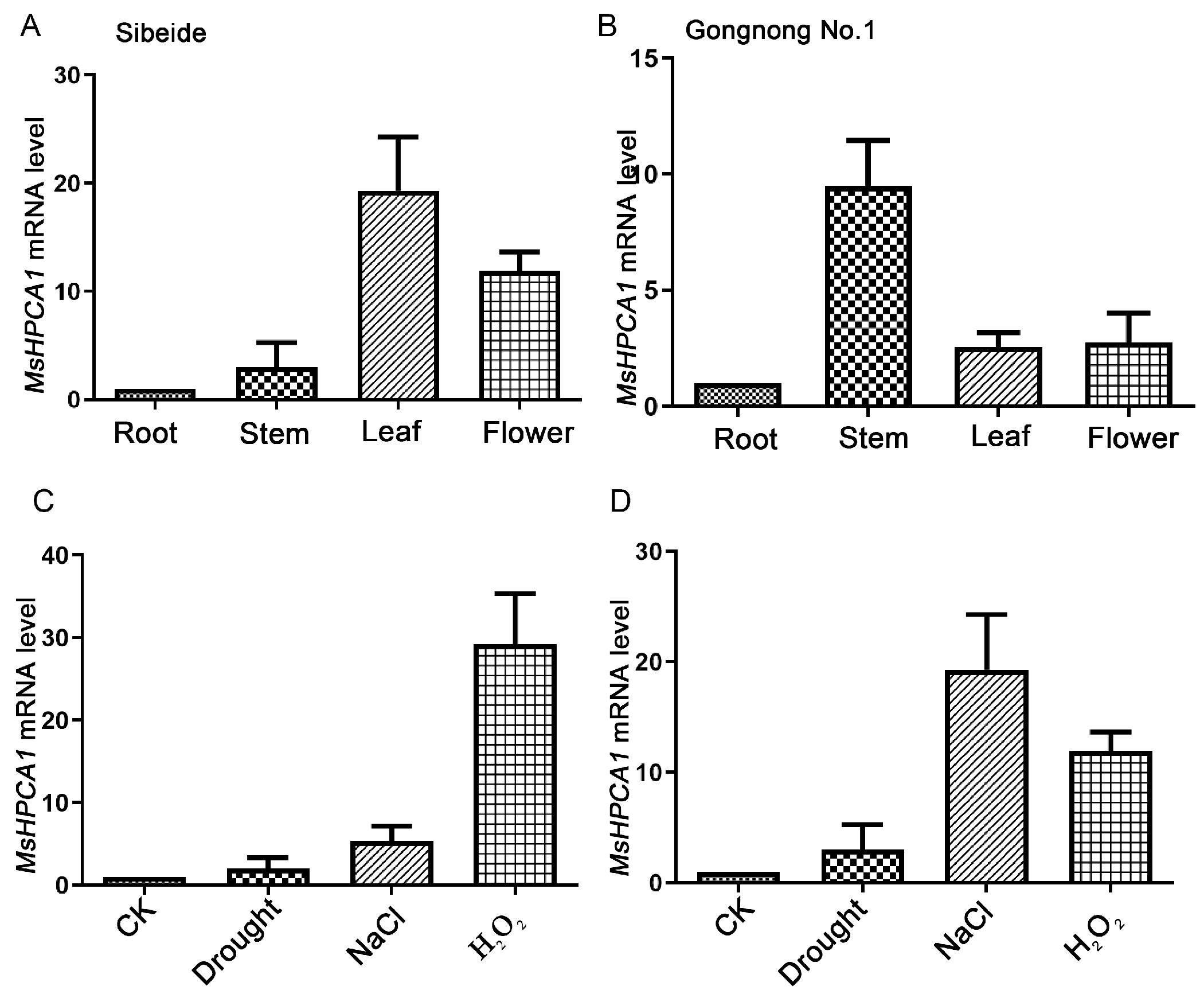
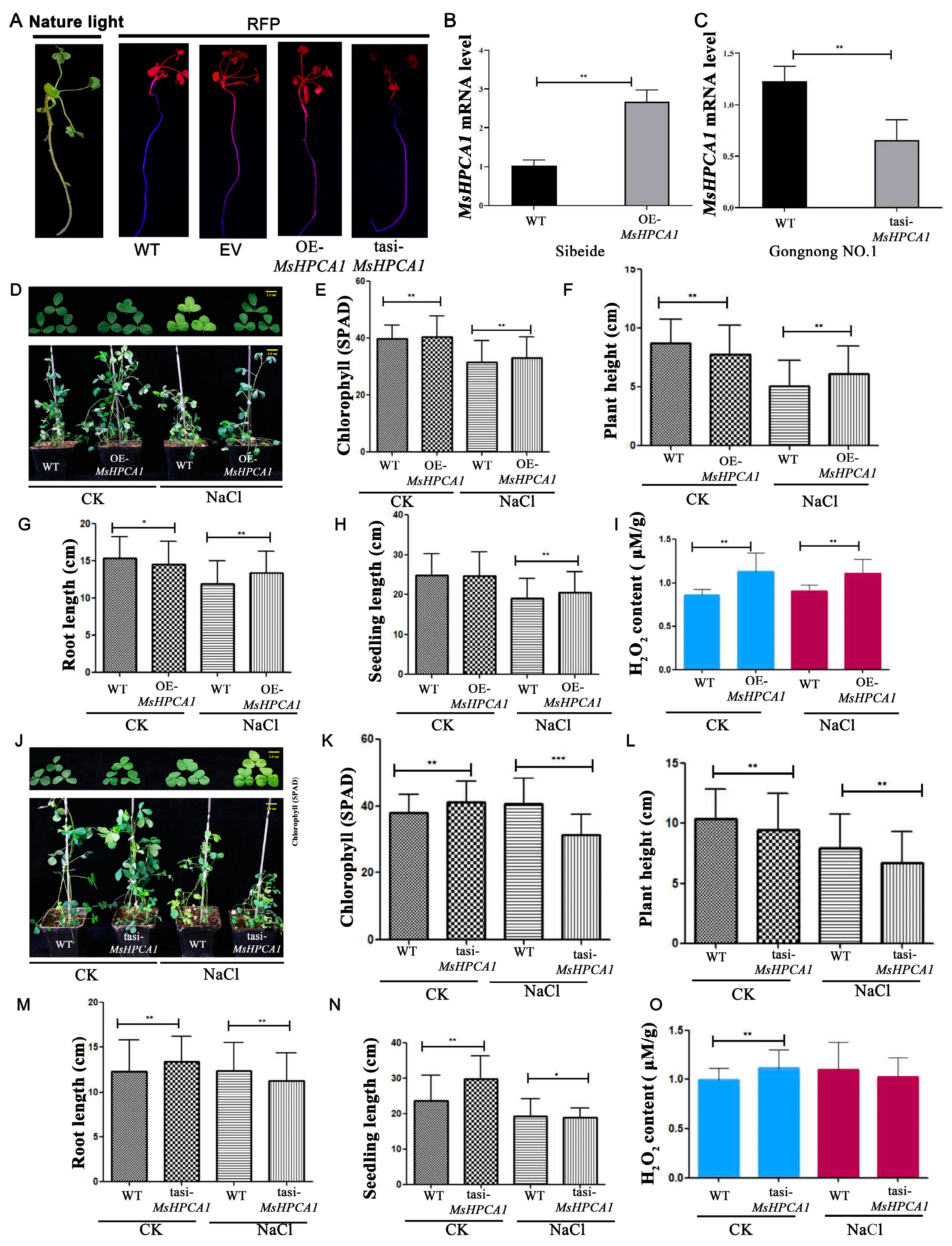
2.5. Expression Analysis of MsHPCA1
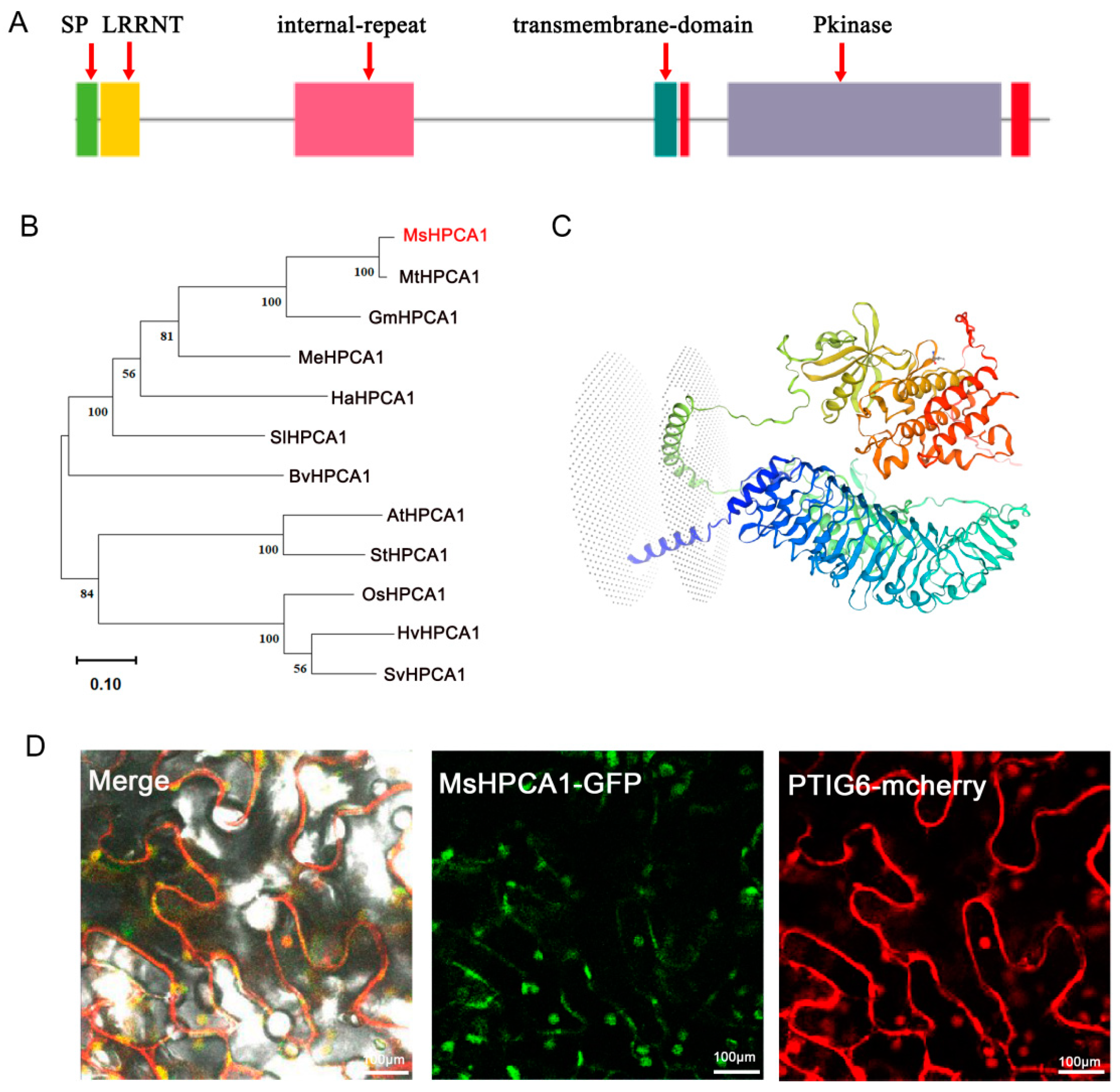
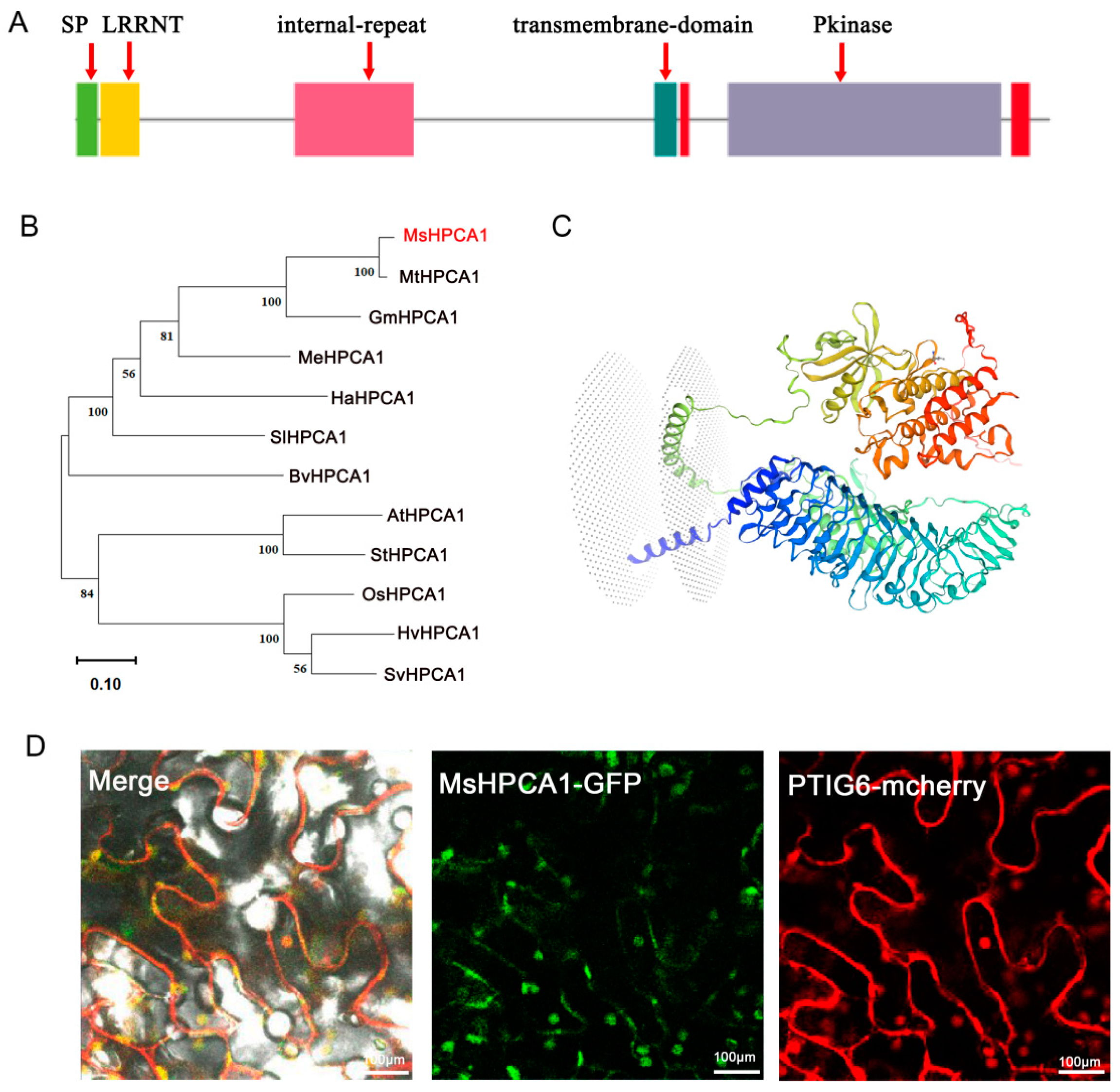
2.6. Subcellular Localization of MsHPCA1
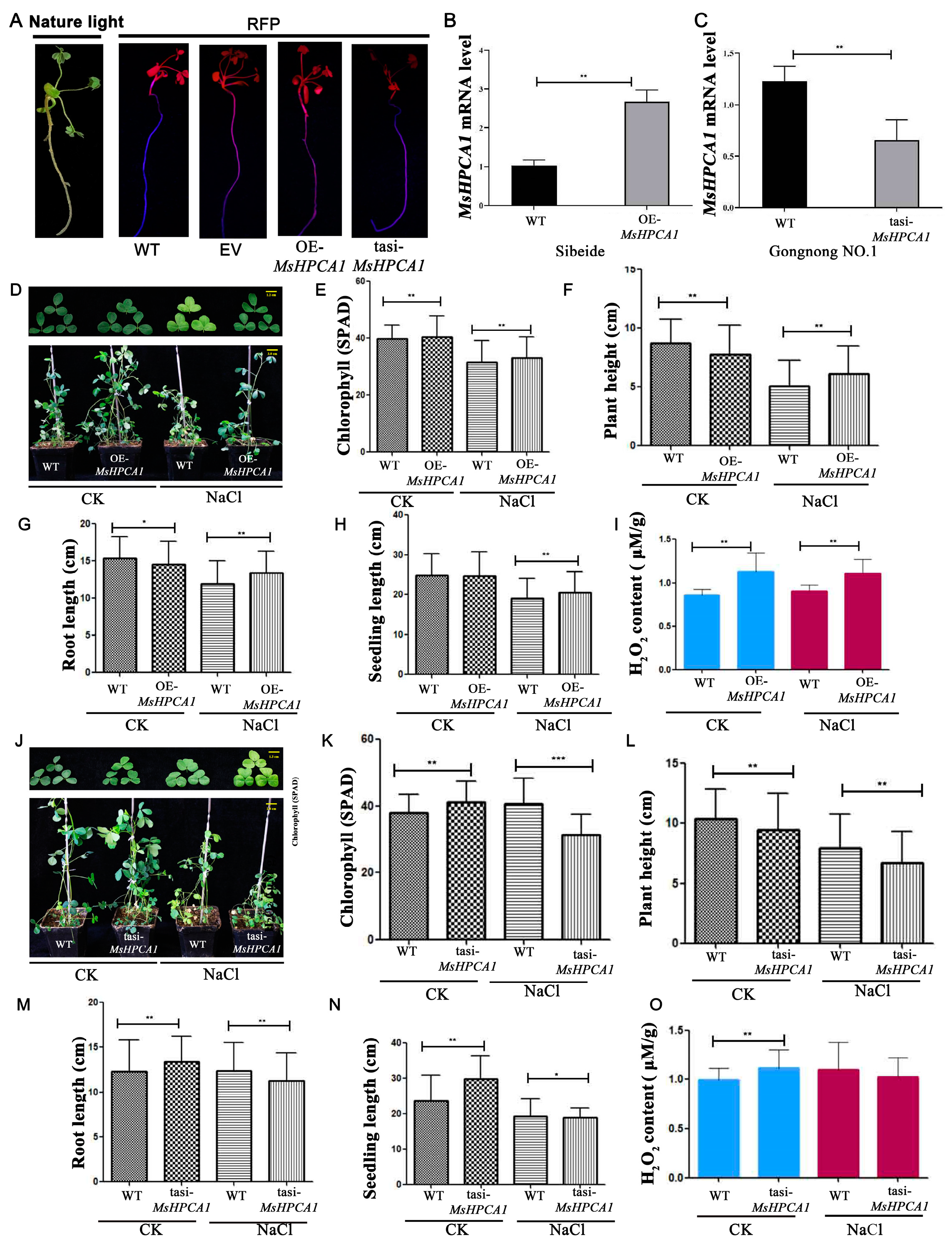
2.7. Transgenic Analysis of MsHPCA1 Function
3. Discussion
3.1. Physiological Responses of Seedlings in Two Different Types of Alfalfa to Salt-Stress
3.2. Effect of Salinity Stress on Carbohydrate and Energy Metabolism
3.3. Role of Stress-Associated Family and TF-Encoding Genes in Salt-Stress
3.4. Differentially Expressed Genes in Response to Salt-Stress
3.5. MsHPCA1 Might Be a H2O2 Receptor in Alfalfa and Is Involved in Stress Responses
4. Materials and Methods
4.1. Plant Materials Subjected to Salt Treatments
4.2. Analysis of Physiological Parameters
4.3. Analysis of RNA-Seq Data, Assembly of Sequences, and Annotation of Genes
4.4. Identification of Differentially Expressed Genes
4.5. Quantitative Real-Time PCR Analysis
4.6. Statistical Analysis
4.7. Development of HPCA1 Overexpression and RNAi Alfalfa Lines
4.8. The Relative Expression Level of MsHPCA1
4.9. The Subcellular Localization of MsHPCA1
4.10. Accession Numbers
4.11. Bioinformatics Analysis of MsHPCA1 Gene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burity, H.A.; Ta, T.C.; Faris, M.A.; Coulman, B.E. Estimation of nitrogen fixation and transfer from alfalfa to associated grasses in mixed swards under field conditions. Plant Soil. 1989, 114, 249–255. [Google Scholar] [CrossRef]
- Yang, S.; Zu, Y.Q.; Li, B.; Bi, Y.F.; Jia, L.; He, Y.M.; Li, Y. Response and intraspecific differences in nitrogen metabolism of alfalfa (Medicago sativa L.) under cadmium stress. Chemosphere 2019, 220, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Artiola, J.F.; Walworth, J.L.; Musil, S.A.; Crimmins, M.A. Soil and Land Pollution. In Environmental and Pollution Science, 3rd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 219–235. [Google Scholar]
- Farissi, M.; Faghire, M.; Bargaz, A.; Bouizgaren, A.; Makoudi, B.; Ghoulam, C. Growth, nutrients concentrations, and enzymes involved in plants nutrition of alfalfa populations under saline conditions. J. Agric. Sci. Technol. 2014, 16, 301–314. [Google Scholar]
- Guo, J.Y.; Shi, G.Y.; Guo, X.Y.; Zhang, L.W.; Xu, W.Y.; Wang, Y.M.; Su, Z.; Hua, J.P. Transcriptome analysis reveals that distinct metabolic pathways operate in salt-tolerant and salt-sensitive upland cotton varieties subjected to salinity stress. Plant Sci. 2015, 238, 33–45. [Google Scholar] [CrossRef]
- Epstein, E.; Norlyn, J.D.; Rush, D.W.; Kingsbury, R.W.; Kelley, D.B.; Cunningham, G.A.; Wrona, A.F. Saline culture of crops: A genetic approach. Science 1980, 210, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Manaa, A.; Ahmed, H.B.; Valot, B.; Bouchet, J.P.; Aschi-Smiti, S.; Causse, M.; Faurobert, M. Salt and genotype impact on plant physiology and root proteome variations in tomato. J. Exp. Bot. 2011, 62, 2797–2813. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. The roles of ROS and ABA in systemic acquired acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Zhao, Z.; Li, Y.; Zhou, K.K.; Su, L.; Zhou, Q.Y. Root transcriptome sequencing and differentially expressed drought-responsive genes in the Platycladus orientalis (L.). Tree Genet. Genomes 2016, 12, 79. [Google Scholar] [CrossRef]
- Wu, J.Z.; Zhao, Q.; Sun, D.Q.; Wu, G.W.; Zhang, L.Y.; Yuan, H.M.; Yu, Y.; Zhang, S.Q.; Yang, X.; Li, Z.G.; et al. Transcriptome analysis of flax (Linum usitatissimum L.) undergoing osmotic stress. Ind. Crops Prod. 2018, 116, 215–223. [Google Scholar] [CrossRef]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through Selenium supplementation. Int. J. Mol. Sci. 2019, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Kumar, I.S.; Rao, S.R.; Vardhini, B.V. Role of phytohormones during salt stress tolerance in plants. Curr. Trends Biotechnol. Pharm. 2015, 9, 334–343. [Google Scholar]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of jasmonic acid in plant regulation and response to abiotic stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Diao, P.F.; Kong, L.Q.; Yu, R.N.; Zhang, M.; Zuo, T.T.; Fan, Y.Y.; Niu, Y.D.; Yan, F.; Wuriyanghan, H. Ethylene Enhances Seed Germination and Seedling Growth under Salinity by Reducing Oxidative Stress and Promoting Chlorophyll Content via ETR2 Pathway. Front. Plant Sci. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Z.; Zhang, H.; Song, C.P.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.N.; Zuo, T.T.; Diao, P.F.; Fu, J.B.; Fan, Y.Y.; Wang, Y.; Zhao, Q.Q.; Ma, X.S.; Lu, W.T.; Li, A.G.; et al. Melatonin enhances seed germination and seedling growth of Medicago sativa under salinity via a putative melatonin receptor MsPMTR1. Front. Plant Sci. 2021, 12, 702875. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Mueller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Yeo, A.R. Salinity resistance: Physiologies and prices. Physiol. Plantarum. 1983, 58, 214–222. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Song, J.; Wang, B.S. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Zhang, F.; Zhang, F.; Wang, W.S.; Zhou, Y.L.; Fu, B.Y.; Li, Z.K. Comparative transcriptome sequencing of tolerant rice introgression line and its parents in response to drought stress. BMC Genom. 2014, 15, 1026. [Google Scholar] [CrossRef] [PubMed]
- Diray-Arce, J.; Clement, M.; Gul, B.; Khan, M.A.; Nielsen, B.L. Transcriptome assembly, profiling and differential gene expression analysis of the halophyte Suaeda fruticosa provides insights into salt tolerance. BMC Genom. 2015, 16, 353. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.X.; Gao, X.X.; Fu, J.Q.; Zhou, J.H.; Wu, X.F. Metabolic response of maize (Zea mays L.) plants to combined drought and salt stress. Plant Soil 2015, 388, 99–117. [Google Scholar] [CrossRef]
- Amirbakhtiar, N.; Ismaili, A.; Ghaffari, M.R.; Mansuri, R.M.; Sanjari, S.; Shobbar, Z.S. Transcriptome analysis of bread wheat leaves in response to salt stress. PLoS ONE 2021, 16, e0254189. [Google Scholar] [CrossRef]
- Zhang, H.X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765–768. [Google Scholar] [CrossRef]
- Xu, C.S.; Xia, C.; Xia, Z.Q.; Zhou, X.J.; Huang, J.; Huang, Z.Q.; Liu, Y.; Jiang, Y.W.; Casteel, S.; Zhang, C.K. Physiological and transcriptomic responses of reproductive stage soybean to drought stress. Plant Cell Rep. 2018, 37, 1611–1624. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.C.; Sun, Y.; Yang, Q.C.; Chao, Y.H.; Kang, J.M.; Jin, H.; Li, Y.; Margaret, G. Screening of genes induced by salt stress from alfalfa. Mol. Biol. Rep. 2010, 37, 745–753. [Google Scholar] [CrossRef]
- Xiong, X.; Wei, Y.Q.; Chen, J.H.; Liu, N.; Zhang, Y.J. Transcriptome analysis of genes and pathways associated with salt tolerance in alfalfa under non-uniform salt stress. Plant Physiol. Biochem. 2020, 151, 323–333. [Google Scholar] [CrossRef]
- An, Y.M.; Song, L.L.; Liu, Y.R.; Shu, Y.J.; Guo, C.H. De Novo Transcriptional Analysis of Alfalfa in Response to Saline-Alkaline Stress. Front. Plant Sci. 2016, 7, 931. [Google Scholar] [CrossRef]
- Postnikova, O.A.; Shao, J.; Nemchinov, L.G. Analysis of the alfalfa root transcriptome in response to salinity stress. Plant Cell Physiol. 2013, 54, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.T.; Xu, Y.X.; Hettenhausen, C.; Lu, C.K.; Shen, G.J.; Zhang, C.P.; Li, J.; Song, J.; Lin, H.H.; Wu, J.Q. Comparative analysis of alfalfa (Medicago sativa L.) leaf transcriptomes reveals genotype-specific salt tolerance mechanisms. BMC Plant Biol. 2018, 18, 35. [Google Scholar] [CrossRef]
- Wu, F.H.; Chi, Y.; Jiang, Z.H.; Xu, Y.Y.; Pei, Z.M. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Ma, M.S.; Sun, Y.M.; Lu, P.; Shi, H.F.; Guo, Z.F.; Zhu, H.F. Comparative Physiological and Transcriptome Profiles Uncover Salt Tolerance Mechanisms in Alfalfa. Front. Plant Sci. 2022, 13, 931619. [Google Scholar] [CrossRef] [PubMed]
- Pommerrenig, B.; Papini-Terzi, F.S.; Sauer, N. Differential Regulation of Sorbitol and Sucrose Loading into the Phloem of Plantago major in Response to Salt Stress. Plant Physiol. 2007, 144, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Boriboonkaset, T.; Theerawitaya, C.; Yamada, N.; Pichakum, A.; Supaibulwatana, K.; Cha-Um, S.; Takabe, T.; Kirdmanee, K. Regulation of some carbohydrate metabolism-related genes, starch and soluble sugar contents, photosynthetic activities and yield attributes of two contrasting rice genotypes subjected to salt stress. Protoplasma 2013, 250, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Blumwald, E. Engineering salt tolerance in plants. Curr. Opin. Biotechnol. 2002, 13, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Kazem, H.A.; Chaichan, M.T.; Al-Waeli, A.H.; Sopian, K. A review of dust accumulation and cleaning methods for solar photovoltaic systems. J. Clean. Prod. 2020, 276, 123187. [Google Scholar] [CrossRef]
- Lee, S.K.; Jeon, J.S. Review: Crucial role of inorganic pyrophosphate in integrating carbon metabolism from sucrose breakdown to starch synthesis in rice endosperm. Plant Sci. 2020, 298, 110572. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Blanco-Herrera, F.; Moreno, A.A.; Tapia, R.; Reyes, F.; Araya, M.; Alessio, C.D.; Parodi, A.; Orellana, A. The UDP-glucose: Glycoprotein glucosyltransferase (UGGT), a key enzyme in ER quality control, plays a significant role in plant growth as well as biotic and abiotic stress in Arabidopsis thaliana. BMC Plant Biol. 2015, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, X.K.; Liu, X.; Yang, X.Q.; Li, Y.J.; Hou, B.K. Rice glycosyltransferase gene UGT2 functions in salt stress tolerance under the regulation of bZIP23 transcription factor. Plant Cell Rep. 2023, 42, 17–28. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Srivastava, S.; Penna, S.; D’Souza, S.F. Thiourea orchestrates regulation of redox state and antioxidant responses to reduce the NaCl-induced oxidative damage in Indian mustard (Brassica juncea (L.) Czern.). Plant Physiol. Biochem. 2011, 49, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Effect of NaCl stress on H2O2 metabolism in rice leaves. Plant Growth Regul. 2000, 30, 151–155. [Google Scholar] [CrossRef]
- Liu, S.K.; Cheng, Y.X.; Zhang, X.X.; Guan, Q.J.; Nishiuchi, S.; Hase, K.; Takano, T. Expression of an NADP-malic enzyme gene in rice (Oryza sativa L.) is induced by environmental stresses; over-expression of the gene in Arabidopsis confers salt and osmotic stress tolerance. Plant Mol. Biol. 2007, 64, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, R.; Wiebel, F.F.; Flessau, A.; Rytka, J.; Beyer, A.; Fröhlich, K.U.; Kunau, W.H. PAS1, a yeast gene required for peroxisome biogenesis, encodes a member of a novel family of putative ATPases. Cell 1991, 64, 499–510. [Google Scholar] [CrossRef]
- Xia, Z.L.; Wei, Y.Y.; Sun, K.L.; Wu, J.Y.; Wang, Y.X.; Wu, K. The maize AAA-type protein SKD1 confers enhanced salt and drought stress tolerance in transgenic tobacco by interacting with Lyst-interacting protein 5. PLoS ONE 2013, 8, e69787. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Wang, T.Z.; Zhang, D.; Chen, L.; Wang, J.; Zhang, W.H. Genome-wide analysis of the Glutathione S-Transferase family in wild Medicago ruthenica and drought-tolerant breeding application of MruGSTU39 gene in cultivated alfalfa. Theor. Appl. Genet. 2022, 135, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Sun, H.; Huang, J.; Zhang, H.S. Functional analysis of a novel Cys2/His2-type zinc finger protein involved in salt tolerance in rice. J. Exp. Bot. 2010, 61, 2807–2818. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Lee, I.H.; Nou, I.S.; Lee, K.D.; Rashotte, A.M.; Kang, K.K. BrRZFP1 a Brassica rapa C3HC4-type RING zinc finger protein involved in cold, salt and dehydration stress. Plant Biol. 2013, 15, 274–283. [Google Scholar] [CrossRef]
- Gupta, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Abiotic stresses cause differential regulation of alternative splice forms of GATA transcription factor in rice. Front. Plant Sci. 2017, 8, 1944. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.H.; Wu, T.T.; Zhao, X.; Wang, Z.Q.; Chen, Y. Comparative physiological and full-length transcriptome analyses reveal the molecular mechanism of melatonin-mediated salt tolerance in okra (Abelmoschus esculentus L.). BMC Plant Biol. 2021, 21, 180. [Google Scholar] [CrossRef]
- Wang, Z.R.; Gao, M.; Li, Y.F.; Zhang, J.L.; Su, H.; Cao, M.; Liu, Z.J.; Zhang, X.C.; Zhao, B.; Guo, Y.D.; et al. The transcription factor SlWRKY37 positively regulates jasmonic acid- and dark-induced leaf senescence in tomato. J. Exp. Bot. 2020, 73, 6207–6225. [Google Scholar] [CrossRef]
- Li, W.Y.; Wang, C.; Shi, H.H.; Wang, B.; Wang, J.X.; Liu, Y.S.; Ma, J.Y.; Tian, S.Y.; Zhang, Y.W. Genome-wide analysis of ethylene-response factor family in adzuki bean and functional determination of VaERF3 under saline-alkaline stress. Plant Physiol. Biochem. 2020, 147, 215–222. [Google Scholar] [CrossRef]
- Vannini, C.; Iriti, M.; Bracale, M.; Locatelli, F.; Faoro, F.; Croce, P.; Pirona, R.; Maro, A.D.; Coraggio, I.; Genga, A. The ectopic expression of the rice Osmyb4 gene in Arabidopsis increases tolerance to abiotic, environmental and biotic stresses. Physiol. Mol. Plant Pathol. 2006, 69, 26–42. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.H.; Cai, H.Y.; Guo, M.L.; Chai, M.N.; She, Z.Y.; Ye, L.; Cheng, Y.; Wang, B.R.; Qin, Y. The bZIP transcription factor GmbZIP15 negatively regulates salt- and drought-stress responses in soybean. Int. J. Mol. Sci. 2020, 21, 7778. [Google Scholar] [CrossRef]
- Liu, A.L.; Zou, J.; Liu, C.F.; Zhou, X.Y.; Zhang, X.W.; Luo, G.Y.; Chen, X.B. Over-expression of OsHsfA7 enhanced salt and drought tolerance in transgenic rice. BMB Rep. 2013, 46, 31–36. [Google Scholar] [CrossRef]
- Zou, J.; Liu, C.F.; Liu, A.L.; Zou, D.; Chen, X.B. Overexpression of OsHsp17.0 and OsHsp23.7 enhances drought and salt tolerance in rice. J. Plant Physiol. 2012, 169, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.C.; Liu, W.Q.; Qiu, L.Y.; Zhang, S.M.; Ma, L.; Zhang, H. Overexpression of glutathione S-transferase gene increases salt tolerance of arabidopsis. Russ. J. Plant Physiol. 2010, 57, 233–240. [Google Scholar] [CrossRef]
- Jin, T.; Sun, Y.Y.; Zhao, R.R.; Shan, Z.; Gai, J.Y.; Li, Y. Overexpression of Peroxidase Gene GsPRX9 Confers Salt Tolerance in Soybean. Int. J. Mol. Sci. 2019, 20, 3745. [Google Scholar] [CrossRef]
- Wang, T.Z.; Zhang, J.L.; Tian, Q.Y.; Zhao, M.G.; Zhang, W.H. A Medicago truncatula EF-hand family gene, MtCaMP1, is involved in drought and salt stress tolerance. PLoS ONE 2013, 8, e58952. [Google Scholar] [CrossRef]
- Battaglia, M.; Covarrubias, A.A. Late Embryogenesis Abundant (LEA) proteins in legumes. Front. Plant Sci. 2013, 4, 190. [Google Scholar] [CrossRef]
- Fichman, Y.; Zandalinas, S.I.; Peck, S.; Luan, S.; Mittler, R. HPCA1 is required for systemic reactive oxygen species and calcium cell-to-cell signaling and plant acclimation to stress. Plant Cell 2022, 34, 4453–4471. [Google Scholar] [CrossRef] [PubMed]
- Laohavisit, A.; Wakatake, T.; Ishihama, N.; Mulvey, H.; Takizawa, K.; Suzuki, T.; Shirasu, K. Quinone perception in plants via leucine-rich-repeat receptor-like kinases. Nature 2020, 587, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, S.B.; Zhang, H.; Li, Y.H.; Niu, L.J.; Zhang, J.H.; Wang, W. Signaling Transduction of ABA, ROS, and Ca2+ in Plant Stomatal Closure in Response to Drought. Int. J. Mol. Sci. 2022, 23, 14824. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- He, F.; Wei, C.; Zhang, Y.X.; Long, R.C.; Li, M.N.; Wang, Z.; Yang, Q.C.; Kang, J.M.; Chen, L. Genome-wide association analysis coupled with transcriptome analysis reveals candidate genes related to salt stress in alfalfa (Medicago sativa L.). Front. Plant Sci. 2022, 3, 826584. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Q.; Yu, R.; Ma, X.; Wuriyanghan, H.; Yan, F. Transcriptome Analysis for Salt-Responsive Genes in Two Different Alfalfa (Medicago sativa L.) Cultivars and Functional Analysis of MsHPCA1. Plants 2024, 13, 1073. https://doi.org/10.3390/plants13081073
Gao Q, Yu R, Ma X, Wuriyanghan H, Yan F. Transcriptome Analysis for Salt-Responsive Genes in Two Different Alfalfa (Medicago sativa L.) Cultivars and Functional Analysis of MsHPCA1. Plants. 2024; 13(8):1073. https://doi.org/10.3390/plants13081073
Chicago/Turabian StyleGao, Qican, Ruonan Yu, Xuesong Ma, Hada Wuriyanghan, and Fang Yan. 2024. "Transcriptome Analysis for Salt-Responsive Genes in Two Different Alfalfa (Medicago sativa L.) Cultivars and Functional Analysis of MsHPCA1" Plants 13, no. 8: 1073. https://doi.org/10.3390/plants13081073
APA StyleGao, Q., Yu, R., Ma, X., Wuriyanghan, H., & Yan, F. (2024). Transcriptome Analysis for Salt-Responsive Genes in Two Different Alfalfa (Medicago sativa L.) Cultivars and Functional Analysis of MsHPCA1. Plants, 13(8), 1073. https://doi.org/10.3390/plants13081073