
Determination of Morpho-Physiological Traits for Assessing Drought Tolerance in Sugarcane

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
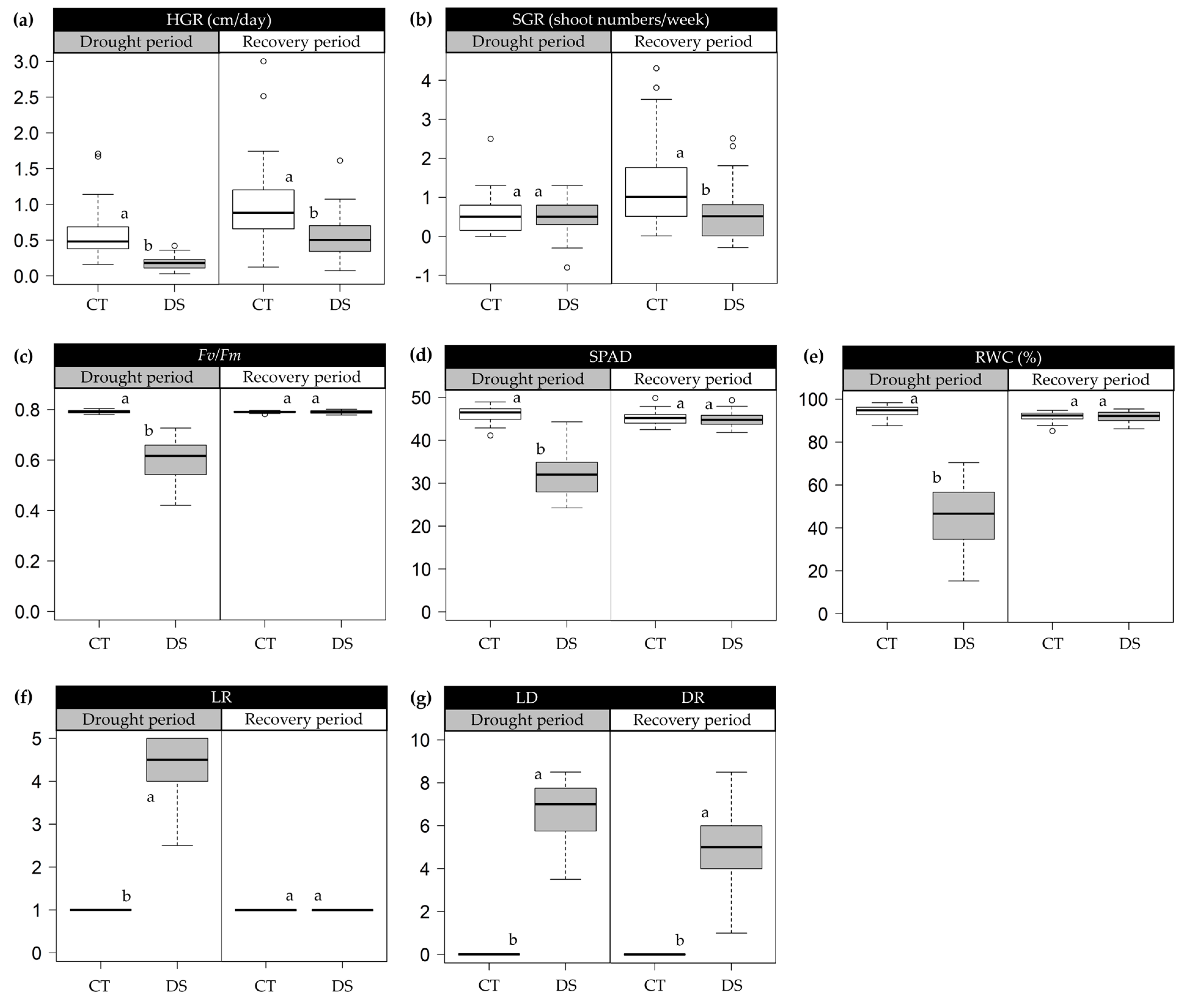
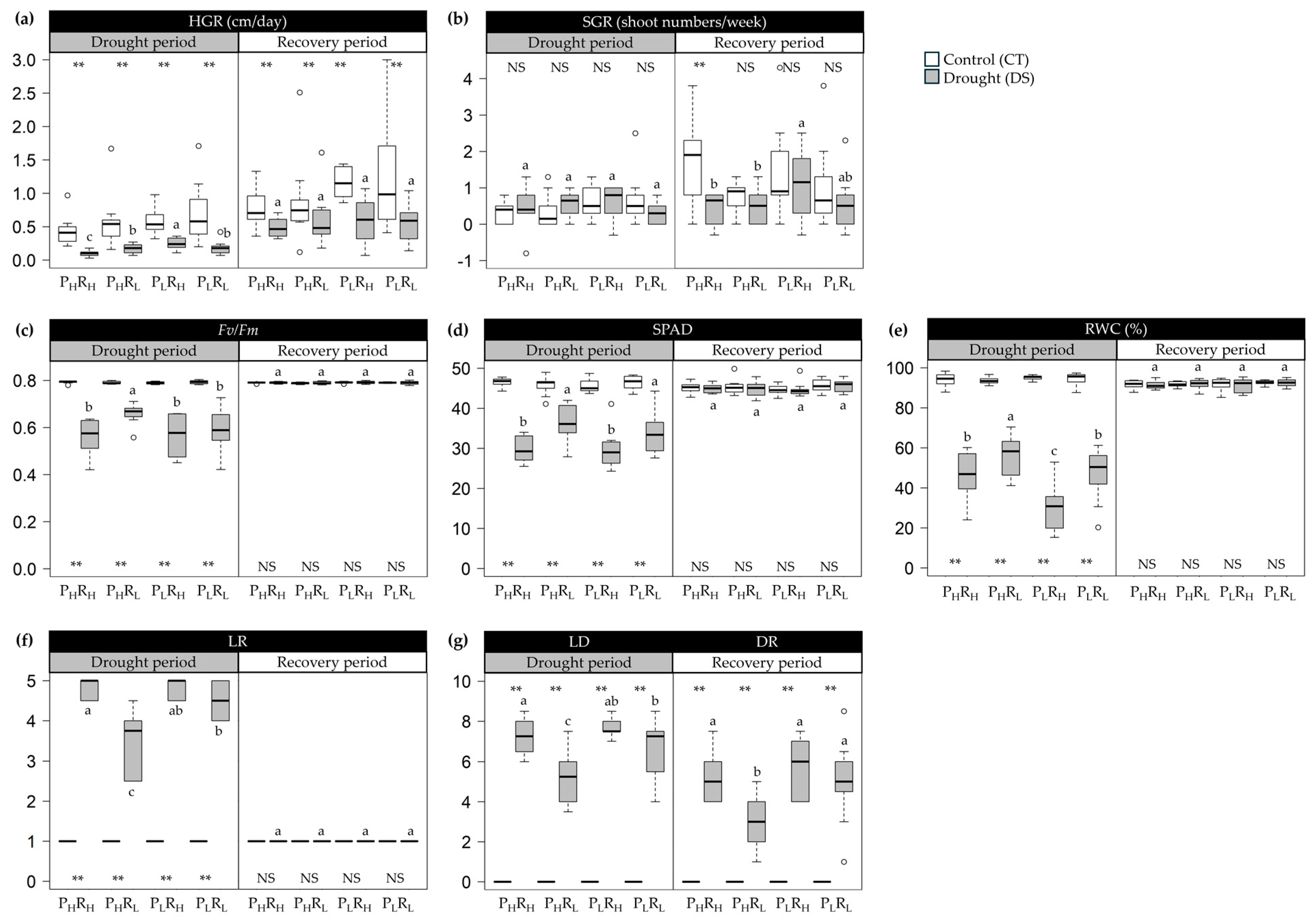
2.1. Morpho-Physiological Variations in Progressive Drought Stress and Recovery
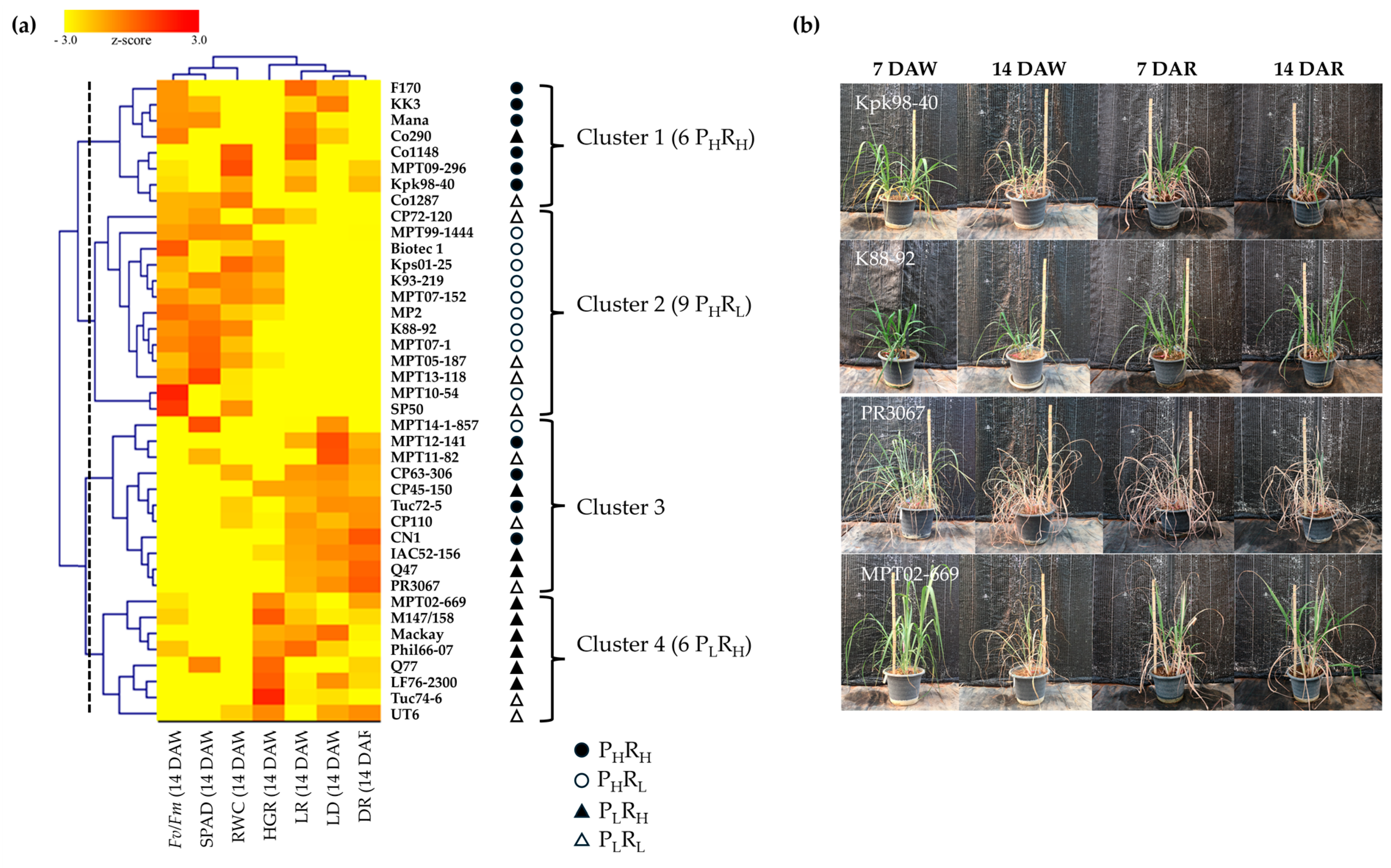
2.2. Hierarchical Clustering of Sugarcane Genotypes Based on Morpho-Physiological Traits
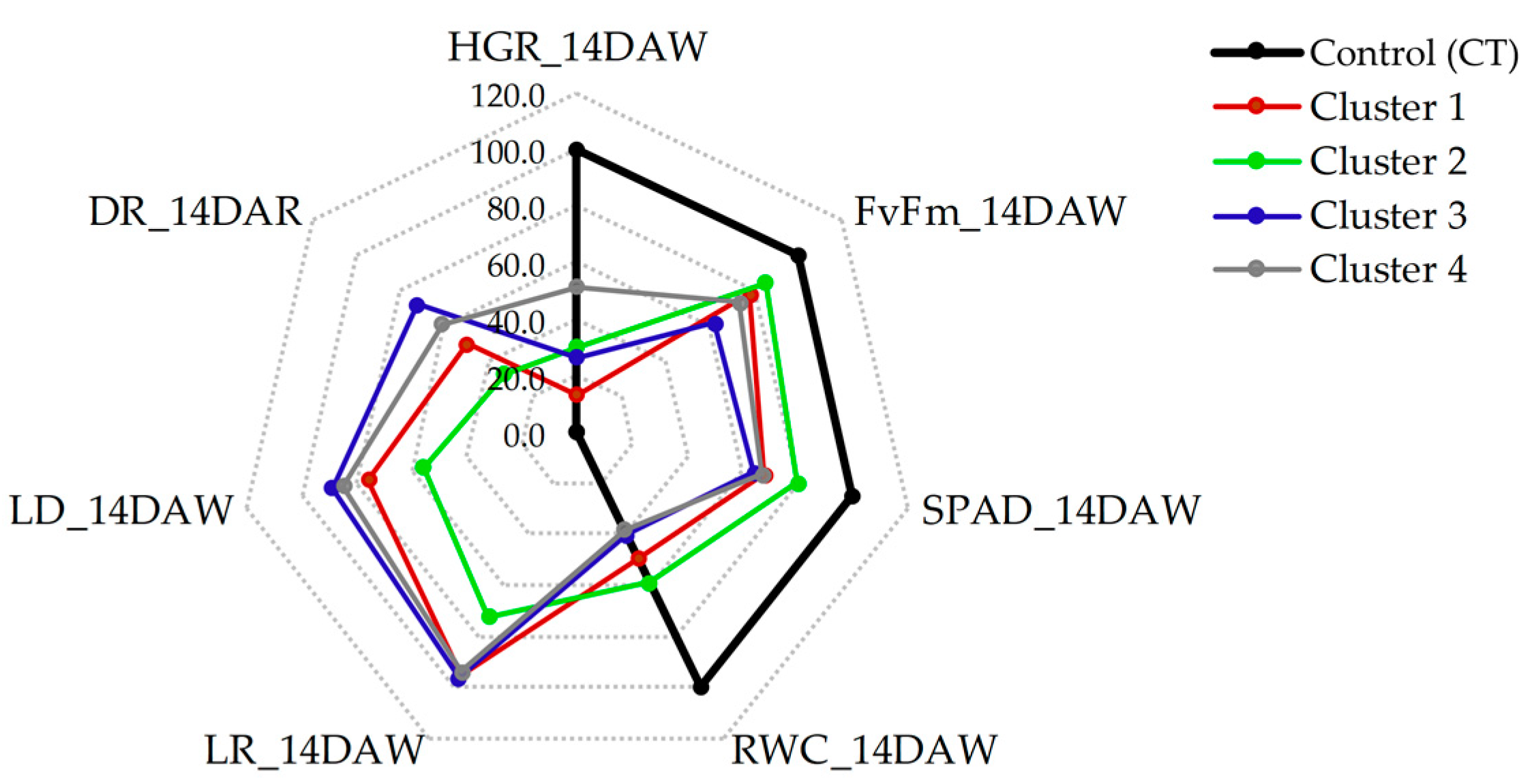
2.3. Variability of the Genotypes in the Clusters
2.4. Principal Component Analysis
2.5. Correlation Analysis
3. Discussion
3.1. Trait Variability under Drought and Recovery Treatments
3.2. Relationships between Genotypes, Traits, and Drought Tolerance
3.3. Morpho-Physiological Traits Related to Stay-Green Phenotype
4. Materials and Methods
4.1. Plant Materials and Stress Treatment
4.2. Soil Moisture Measurement and Meteorological Conditions
4.3. Morpho-Physiological Measurements
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lakshmanan, P.; Robinson, N. Stress Physiology: Abiotic Stresses. In Sugarcane: Physiology, Biochemistry, and Functional Biology, 1st ed.; Paul, H.M., Frederik, C.B., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 411–434. [Google Scholar]
- Kumar, D.; Malik, N.; Sengar, R.S.; Yadav, B.; Singh, A.K.; Yadav, C.L.; Yadav, M.K. Transcriptional regulation in sugarcane under water deficit during formative growth stage. Cytol. Genet. 2021, 55, 598–605. [Google Scholar] [CrossRef]
- Mahadevaiah, C.; Hapase, P.; Sreenivasa, V.; Hapase, R.; Swamy, H.K.M.; Anilkumar, C.; Mohanraj, K.; Hemaprabha, G.; Ram, B. Delineation of genotype × environment interaction for identification of stable genotypes for tillering phase drought stress tolerance in sugarcane. Sci. Rep. 2021, 11, 18649. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.A.O.; Carpentero, A.S.; Santos, P.J.A.; Delfin, E.F. Effects of water regime, genotype, and formative stages on the agro-physiological response of sugarcane (Saccharum officinarum L.) to drought. Plants 2020, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.A.; Casas, D.; Gandia, J.; Delfin, E. Drought impact on sugarcane production. In Agricultural Research Updates; Gorawala, P., Mandhatri, S., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2021; Volume 35, pp. 53–93. [Google Scholar]
- Khonghintaisong, J.; Songsri, P.; Jongrungklang, N. Understanding growth rate patterns among different drought resistant sugarcane cultivars during plant and ratoon crops encountered water deficit at early growth stage under natural field conditions. Agronomy 2021, 11, 2083. [Google Scholar] [CrossRef]
- Ferreira, T.H.S.; Tsunada, M.S.; Bassi, D.; Araújo, P.; Mattiello, L.; Guidelli, G.V.; Righetto, G.L.; Gonçalves, V.R.; Lakshmanan, P.; Menossi, M. Sugarcane water stress tolerance mechanisms and its implications on developing biotechnology solutions. Front. Plant Sci. 2017, 8, 1077. [Google Scholar] [CrossRef] [PubMed]
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding Strategies to Enhance Drought Tolerance in Crops. In Advances in Plant Breeding Strategies: Agronomic, Abiotic and Biotic Stress Traits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 397–445. [Google Scholar]
- Toum, L.; Perez-Borroto, L.S.; Peña-Malavera, A.N.; Luque, C.; Welin, B.; Berenstein, A.; Fernández Do Porto, D.; Vojnov, A.; Castagnaro, A.P.; Pardo, E.M. Selecting putative drought-tolerance markers in two contrasting soybeans. Sci. Rep. 2022, 12, 10872–10888. [Google Scholar] [CrossRef]
- Srivastava, M.K.; Li, C.-N.; Li, Y.-R. Development of sequence characterized amplified region (SCAR) marker for identifying drought tolerant sugarcane genotypes. Aust. J. Crop Sci. 2012, 6, 763–767. [Google Scholar]
- Khaled, K.; El Sherbeny, S.; Abdelhadi, A. R-ISSR as marker assisted selection for drought tolerance In sugarcane. J. Agric. Chem. Biotechnol. 2017, 8, 91–97. [Google Scholar] [CrossRef]
- Adly Khaled, K.; El-Arabi, N.I.; Mahmoud Sabry, N.; El-Sherbiny, S. Sugarcane genotypes assessment under drought condition using amplified fragment length polymorphism. Biotechnology 2018, 17, 120–127. [Google Scholar] [CrossRef]
- Jangpromma, N.; Songsri, P.; Thammasirirak, S.; Jaisil, P. Rapid assessment of chlorophyll content in sugarcane using a SPAD chlorophyll meter across different water stress conditions. Asian J. Plant Sci. 2010, 9, 368–374. [Google Scholar] [CrossRef]
- Silva, M.D.A.; Jifon, J.L.; Sharma, V.; da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of physiological parameters in screening drought tolerance in sugarcane genotypes. Sugar Tech 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Yasir, T.A.; Wasaya, A.; Hussain, M.; Ijaz, M.; Farooq, M.; Farooq, O.; Nawaz, A.; Hu, Y.G. Evaluation of physiological markers for assessing drought tolerance and yield potential in bread wheat. Physiol. Mol. Biol. Plants 2019, 25, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Fried, H.G.; Narayanan, S.; Fallen, B. Evaluation of soybean [Glycine max (L.) Merr.] genotypes for yield, water use efficiency, and root traits. PLoS ONE 2019, 14, e0212700. [Google Scholar] [CrossRef] [PubMed]
- Leanasawat, N.; Kosittrakun, M.; Lontom, W.; Songsri, P. Physiological and agronomic traits of certain sugarcane genotypes grown under field conditions as influenced by early drought stress. Agronomy 2021, 11, 2319. [Google Scholar] [CrossRef]
- Silva, M.; Jifon, J.; Da Silva, J.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Dordas, C.; Papathanasiou, F.; Lithourgidis, A.; Petrevska, J.-K.; Papadopoulos, I.; Pankou, C.; Gekas, F.; Ninou, E.; Mylonas, I.; Sistanis, I.; et al. Evaluation of physiological characteristics as selection criteria for drought tolerance in maize inbred lines and their hybrids. Maydica 2018, 63, 1–14. [Google Scholar]
- da Silva, P.P.; Soares, L.; da Costa, J.G.; da Silva Viana, L.; de Andrade, J.C.F.; Gonçalves, E.R.; dos Santos, J.M.; de Souza Barbosa, G.V.; Nascimento, V.X.; Todaro, A.R.; et al. Path analysis for selection of drought tolerant sugarcane genotypes through physiological components. Ind. Crops Prod. 2012, 37, 11–19. [Google Scholar] [CrossRef]
- Benešová, M.; Holá, D.; Fischer, L.; Jedelský, P.L.; Hnilička, F.; Wilhelmová, N.; Rothová, O.; Kočová, M.; Procházková, D.; Honnerová, J.; et al. The physiology and proteomics of drought tolerance in maize: Early stomatal closure as a cause of lower tolerance to short-term dehydration? PLoS ONE 2012, 7, e38017. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Ougham, H. The stay-green trait. J. Exp. Bot. 2014, 65, 3889–3900. [Google Scholar] [CrossRef]
- Kumar, R.; Krishna, H.; Barman, D.; Ghimire, O.; Shekharappa, G.; Singh, P.; Chinnusamy, V.; Padaria, J.; Arora, A. Stay-green trait serves as yield stability attribute under combined heat and drought stress in wheat (Triticum aestivum L.). Plant Growth Regul. 2021, 96, 67–78. [Google Scholar] [CrossRef]
- Khonghintaisong, J.; Songsri, P.; Toomsan, B.; Jongrungklang, N. Rooting and physiological trait responses to early drought stress of sugarcane cultivars. Sugar Tech 2018, 20, 396–406. [Google Scholar] [CrossRef]
- Chumphu, S.; Jongrungklang, N.; Songsri, P. Association of physiological responses and root distribution patterns of ratooning ability and yield of the second ratoon cane in sugarcane elite clones. Agronomy 2019, 9, 200–217. [Google Scholar] [CrossRef]
- Inman-Bamber, N.G.; Lakshmanan, P.; Park, S. Sugarcane for water-limited environments: Theoretical assessment of suitable traits. Field Crops Res. 2012, 134, 95–104. [Google Scholar] [CrossRef]
- Madhav, T.; Madhu, B.G.S.; Mede, V.; Naik, C. Study on Root Characteristics of Sugarcane (Saccharum officinarum) Genotypes for Moisture Stress. Int. J. Plant Soil Sci. 2017, 18, 1–4. [Google Scholar] [CrossRef]
- De Almeida Silva, M.; Jifon, J.L.; Da Silva, J.A.G.; Dos Santos, C.M.; Sharma, V. Relationships between physiological traits and productivity of sugarcane in response to water deficit. J. Agric. Sci. 2014, 152, 104–118. [Google Scholar] [CrossRef]
- Yan, C.; Song, S.; Wang, W.; Wang, C.; Li, H.; Wang, F.; Li, S.; Sun, X. Screening diverse soybean genotypes for drought tolerance by membership function value based on multiple traits and drought-tolerant coefficient of yield. BMC Plant Biol. 2020, 20, 321–335. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Silva, M.; Gonçalves da Silva, J.A.; Enciso, J.; Sharma, V.; Jifon, J. Yield components as indicators of drought tolerance of sugarcane. Sci. Agric. 2008, 65, 620–627. [Google Scholar] [CrossRef]
- Jangpromma, N.; Thammasirirak, S.; Jaisil, P.; Songsri, P. Effects of drought and recovery from drought stress on above ground and root growth, and water use efficiency in sugarcane (Saccharum officinarum L.). Aust. J. Crop Sci. 2012, 6, 1298–1304. [Google Scholar]
- Chapae, C.; Songsri, P.; Gonkhamdee, S.; Jongrungklang, N. Understanding drought responses of sugarcane cultivars controlled under low water potential conditions. Chil. J. Agric. Res. 2020, 80, 370–380. [Google Scholar] [CrossRef]
- Tippayawat, A.; Jogloy, S.; Vorasoot, N.; Songsri, P.; Kimbeng, C.A.; Jifon, J.L.; Janket, A.; Thangthong, N.; Jongrungklang, N. Differential physiological responses to different drought durations among a diverse set of sugarcane genotypes. Agronomy 2023, 13, 2594. [Google Scholar] [CrossRef]
- Jiang, Y.; Yao, Y.; Wang, Y. Physiological response, cell wall components, and gene expression of switchgrass under short-term drought stress and recovery. Crop Sci. 2012, 52, 2718. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Tran, H.; Shan, L.; Kim, J.; Childs, K.; Ervin, E.H.; Frazier, T.; Zhao, B. Assessment of drought tolerance of 49 switchgrass (Panicum virgatum) genotypes using physiological and morphological parameters. Biotechnol. Biofuels 2015, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Khan, A.; Li, M.-J.; Khan, S.; Kashif, M. Early selection of bread wheat genotypes using morphological and photosynthetic attributes conferring drought tolerance. J. Integr. Agric. 2019, 18, 2483–2491. [Google Scholar] [CrossRef]
- Mohi-Ud-Din, M.; Hossain, M.A.; Rohman, M.M.; Uddin, M.N.; Haque, M.S.; Ahmed, J.U.; Hossain, A.; Hassan, M.M.; Mostofa, M.G. Multivariate analysis of morpho-physiological traits reveals differential drought tolerance potential of bread wheat genotypes at the seedling stage. Plants 2021, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Lean, F.L.; Ismail, M.; Berahim, Z.; Rahman, M.; Islam, M. Physiological and molecular characterization of drought responses and screening of drought tolerant rice varieties. Biosci. J. 2015, 31, 709–718. [Google Scholar] [CrossRef]
- Swapna, S.; Shylaraj, K.S. Screening for osmotic stress responses in rice varieties under drought condition. Rice Sci. 2017, 24, 253–263. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef]
- Cominelli, E.; Conti, L.; Tonelli, C.; Galbiati, M. Challenges and perspectives to improve crop drought and salinity tolerance. New Biotechnol. 2013, 30, 355–361. [Google Scholar] [CrossRef]
- Balbaa, M.G.; Osman, H.T.; Kandil, E.E.; Javed, T.; Lamlom, S.F.; Ali, H.M.; Kalaji, H.M.; Wróbel, J.; Telesiñski, A.; Brysiewicz, A.; et al. Determination of morpho-physiological and yield traits of maize inbred lines (Zea mays L.) under optimal and drought stress conditions. Front. Plant Sci. 2022, 13, 959203. [Google Scholar] [CrossRef]
- de Camargo, M.S.; Bezerra, B.K.L.; Holanda, L.A.; Oliveira, A.L.; Vitti, A.C.; Silva, M.A. Silicon fertilization improves physiological responses in sugarcane cultivars grown under water deficit. J. Soil Sci. Plant Nutr. 2019, 19, 81–91. [Google Scholar] [CrossRef]
- Zhao, D.; Irey, M.; LaBorde, C.; Hu, C.-J. Physiological and yield characteristics of 18 sugarcane genotypes grown on a sand soil. Crop Sci. 2019, 59, 2741–2751. [Google Scholar] [CrossRef]
- Sajid, M.; Amjid, M.; Munir, H.; Ahmad, M.; Zulfiqar, U.; Ali, M.F.; Abul Farah, M.; Ahmed, M.A.A.; Artyszak, A. Comparative analysis of growth and physiological responses of sugarcane elite genotypes to water stress and sandy loam soils. Plants 2023, 12, 2759. [Google Scholar] [CrossRef] [PubMed]
- Gholamin, R.; Khayatnezhad, M. The effect of end season drought stress on the chlorophyll content, chlorophyll fluorescence parameters and yield in maize cultivars. Sci. Res. Essays 2011, 6, 5351–5357. [Google Scholar]
- Effendi, R.; Priyanto, S.B.; Aqil, M.; Azrai, M. Drought adaptation level of maize genotypes based on leaf rolling, temperature, relative moisture content, and grain yield parameters. IOP Conf. Ser. Earth Environ. Sci. 2019, 270, 012016. [Google Scholar] [CrossRef]
- Ndikuryayo, C.; Ndayiragije, A.; Kilasi, N.L.; Kusolwa, P. Identification of drought tolerant rice (Oryza Sativa L.) genotypes with Asian and African backgrounds. Plants 2023, 12, 922. [Google Scholar] [CrossRef] [PubMed]
- Hemaprabha, G.; Swapna, S.; Lavanya, D.L.; Sajitha, B.; Venkataramana, S. Evaluation of drought tolerance potential of elite genotypes and progenies of sugarcane (Saccharum sp. hybrids). Sugar Tech 2013, 15, 9–16. [Google Scholar] [CrossRef]
- Devi, K.; Gomathi, R.; Arun Kumar, R.; Manimekalai, R.; Selvi, A. Field tolerance and recovery potential of sugarcane varieties subjected to drought. Indian J. Plant Physiol. 2018, 23, 271–282. [Google Scholar] [CrossRef]
- Mohanraj, K.; Hemaprabha, G.; Vasantha, S. Biomass yield, dry matter partitioning and physiology of commercial and Erianthus introgressed sugarcane clones under contrasting water regimes. Agric. Water Manag. 2021, 255, 107035. [Google Scholar] [CrossRef]
- Dlamini, P.J. Drought stress tolerance mechanisms and breeding effort in sugarcane: A review of progress and constraints in South Africa. Plant Stress 2021, 2, 100027. [Google Scholar] [CrossRef]
- Chatterjee, J.; Dionora, J.; Elmido-Mabilangan, A.; Wanchana, S.; Thakur, V.; Bandyopadhyay, A.; Brar, D.S.; Quick, W.P. The evolutionary basis of naturally diverse rice leaves anatomy. PLoS ONE 2016, 11, e0164532. [Google Scholar] [CrossRef]
- Matschi, S.; Vasquez, M.F.; Bourgault, R.; Steinbach, P.; Chamness, J.; Kaczmar, N.; Gore, M.A.; Molina, I.; Smith, L.G. Structure-function analysis of the maize bulliform cell cuticle and its potential role in dehydration and leaf rolling. Plant Direct 2020, 4, e00282. [Google Scholar] [CrossRef]
- Latif, A.; Ali, N.; Cuixia, P.; Ying, S. Rice Curled Its Leaves Either Adaxially or Abaxially to Combat Drought Stress. Rice Sci. 2023, 30, 405–416. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef]
- Zhang, T.; Kesoju, S.; Greene, S.L.; Fransen, S.; Hu, J.; Yu, L.-X. Genetic diversity and phenotypic variation for drought resistance in alfalfa (Medicago sativa L.) germplasm collected for drought tolerance. Genet. Resour. Crop Evol. 2018, 65, 471–484. [Google Scholar] [CrossRef]
- Getnet, Z.; Husen, A.; Fetene, M.; Yemata, G. Growth, water status, physiological, biochemical and yield response of stay green Sorghum (Sorghum bicolor (L.) Moench) varieties-a field trial under drought-prone area in amhara regional state, Ethiopia. J. Agron. 2015, 14, 188–202. [Google Scholar] [CrossRef]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Abbas, N.; Ashraf, M.; Ahmad, P. Roles of potential plant hormones and transcription factors in controlling leaf senescence and drought tolerance. Protoplasma 2019, 256, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Wirojsirasak, W.; Songsri, P.; Jongrungklang, N.; Tangphatsornruang, S.; Klomsa-ard, P.; Ukoskit, K. A large-scale candidate-gene association mapping for drought tolerance and agronomic traits in sugarcane. Int. J. Mol. Sci. 2023, 24, 12801. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Jaemsaeng, R.; Jantasuriyarat, C.; Thamchaipenet, A. Molecular interaction of 1-aminocyclopropane-1-carboxylate deaminase (ACCD)-producing endophytic Streptomyces sp. GMKU 336 towards salt-stress resistance of Oryza sativa L. cv. KDML105. Sci. Rep. 2018, 8, 1950. [Google Scholar] [CrossRef]
- International Rice Research Institute (IRRI). Standard Evaluation System (SES) for Rice, 5th ed.; International Rice Research Institute (IRRI): Manila, Philippines, 2013. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 28 June 2023).
- Sturn, A.; Quackenbush, J.; Trajanoski, Z. Genesis: Cluster analysis of microarray data. Bioinformatics 2002, 18, 207–208. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
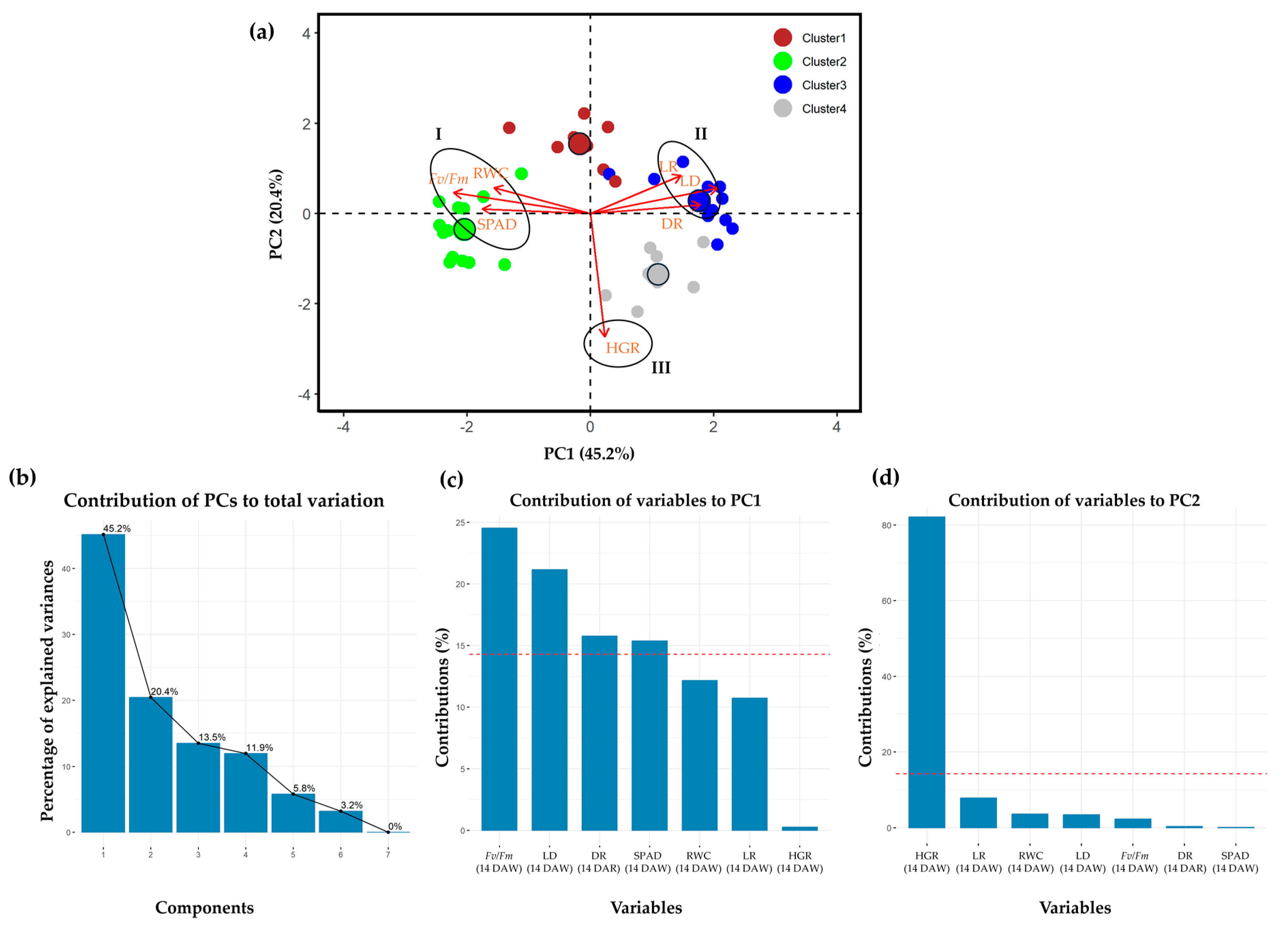{kind=link}
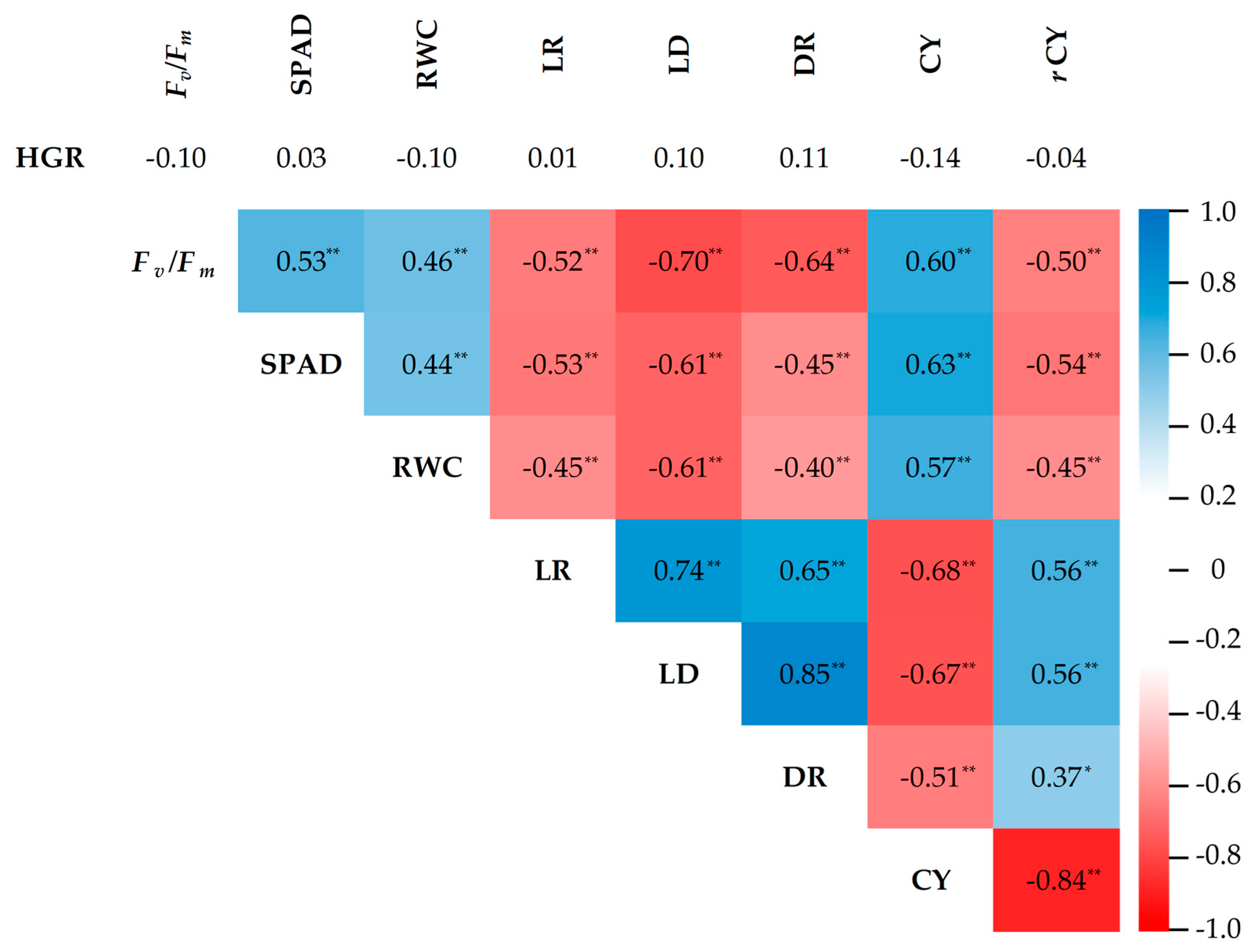{kind=link}
| Cluster | Treatment | Drought Period | Recovery Period | Yield (ton/ha.) | |||||
|---|---|---|---|---|---|---|---|---|---|
| HGR | Fv /Fm | SPAD | RWC | LR | LD | DR | |||
| 1 | CT | 0.54 | 0.792 | 46.7 | 92.91 | 1.0 | 0 | 0 | 121.20 |
| DS | 0.08 c | 0.619 b | 31.3 b | 46.91 ab | 4.8 a | 6.8 b | 4.5 b | 65.27 b | |
| % Change | (-) 85.19 C | (-) 21.84 B | (-) 32.98 B | (-) 49.51 AB | 96.0 A | 75.56 B | 50.0 B | (-) 46.15 B | |
| 2 | CT | 0.53 | 0.792 | 45.7 | 94.31 | 1.0 | 0 | 0 | 122.14 |
| DS | 0.18 b | 0.672 a | 36.7 a | 55.62 a | 3.6 b | 5.0 c | 3.0 c | 106.84 a | |
| % Change | (-) 66.34 AB | (-) 15.15 A | (-) 19.69 A | (-) 41.02 A | 72.0 B | 55.56 C | 33.33 C | (-) 12.53 A | |
| 3 | CT | 0.60 | 0.793 | 46.0 | 94.31 | 1.0 | 0 | 0 | 115.19 |
| DS | 0.15 b | 0.491 c | 29.6 b | 37.76 b | 4.8 a | 8.0 a | 6.5 a | 67.27 b | |
| % Change | (-) 75.0 BC | (-) 38.08 C | (-) 35.65 B | (-) 59.96 B | 96.0 A | 88.89 A | 72.22 A | (-) 41.60 B | |
| 4 | CT | 0.69 | 0.788 | 46.0 | 95.68 | 1.0 | 0 | 0 | 100.98 |
| DS | 0.30 a | 0.582 b | 30.9 b | 36.06 b | 4.7 a | 7.6 a | 5.5 ab | 61.04 b | |
| % Change | (-) 56.52 A | (-) 26.14 B | (-) 32.83 B | (-) 62.31 B | 94.0 A | 84.44 A | 61.11 AB | (-) 39.55 B | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wirojsirasak, W.; Songsri, P.; Jongrungklang, N.; Tangphatsornruang, S.; Klomsa-ard, P.; Ukoskit, K. Determination of Morpho-Physiological Traits for Assessing Drought Tolerance in Sugarcane. Plants 2024, 13, 1072. https://doi.org/10.3390/plants13081072
Wirojsirasak W, Songsri P, Jongrungklang N, Tangphatsornruang S, Klomsa-ard P, Ukoskit K. Determination of Morpho-Physiological Traits for Assessing Drought Tolerance in Sugarcane. Plants. 2024; 13(8):1072. https://doi.org/10.3390/plants13081072
Chicago/Turabian StyleWirojsirasak, Warodom, Patcharin Songsri, Nakorn Jongrungklang, Sithichoke Tangphatsornruang, Peeraya Klomsa-ard, and Kittipat Ukoskit. 2024. "Determination of Morpho-Physiological Traits for Assessing Drought Tolerance in Sugarcane" Plants 13, no. 8: 1072. https://doi.org/10.3390/plants13081072