
Copper Oxide Nanoparticles Induced Growth and Physio-Biochemical Changes in Maize (Zea mays L.) in Saline Soil

 , ,
, ,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
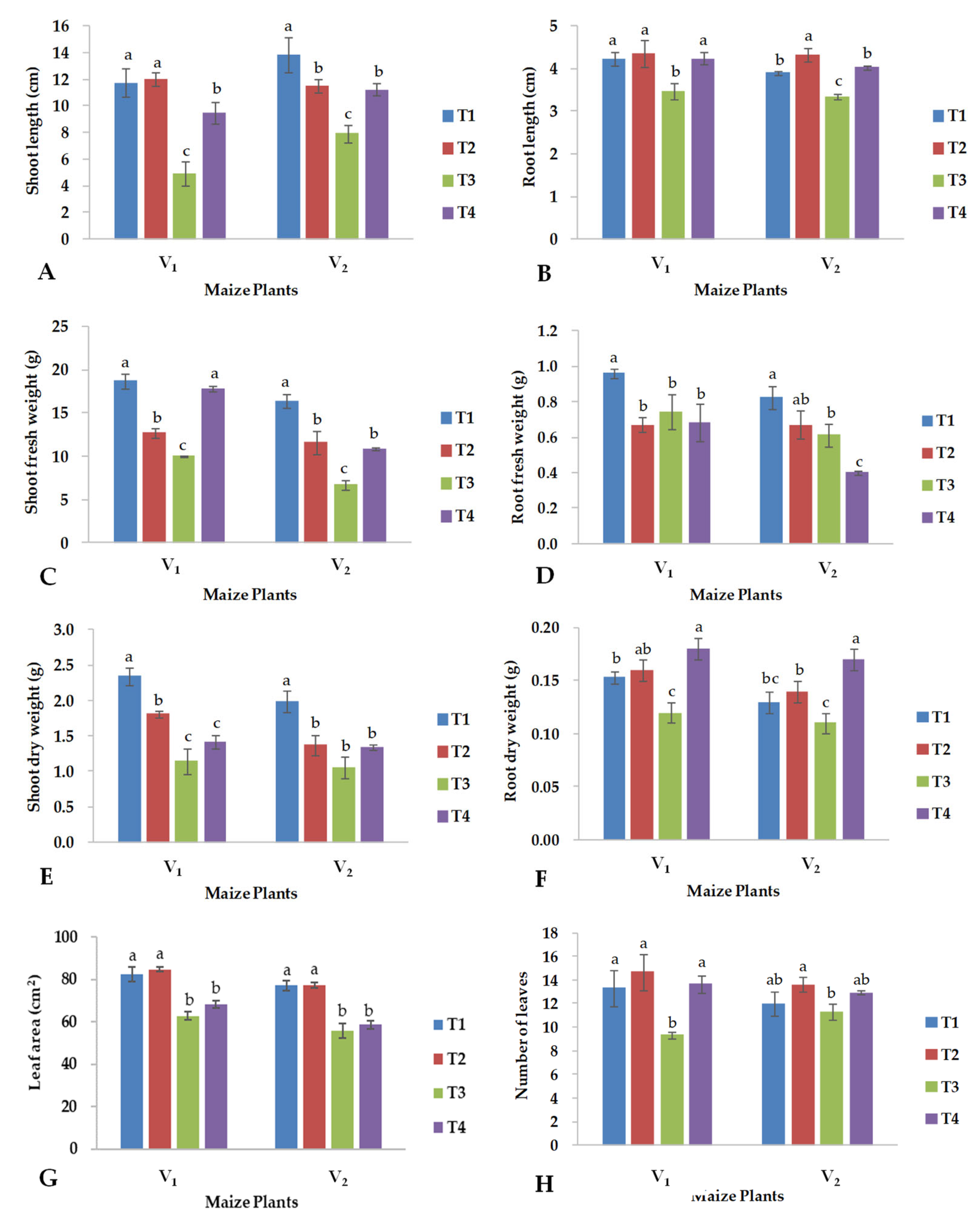
2.1. Growth Parameters
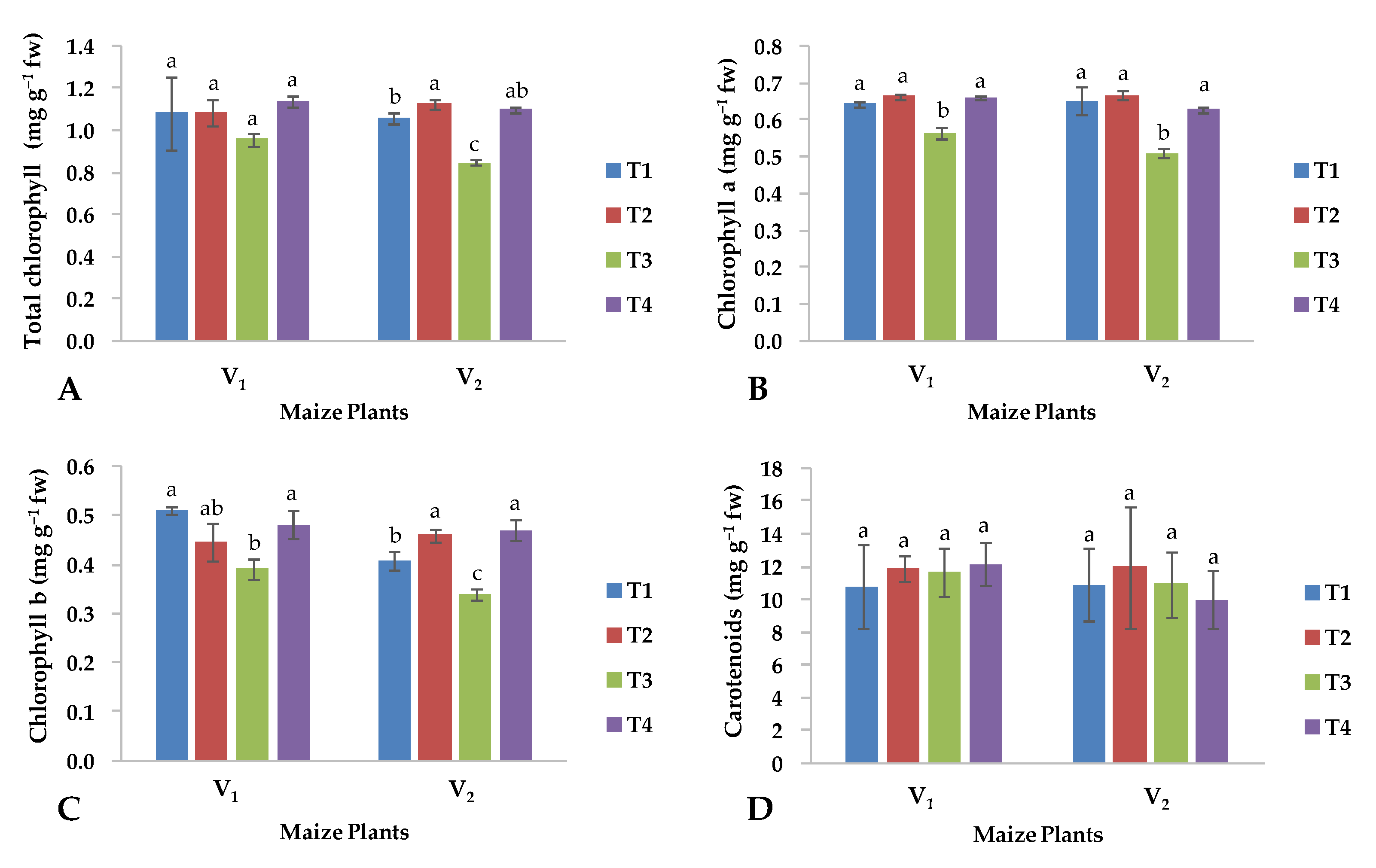
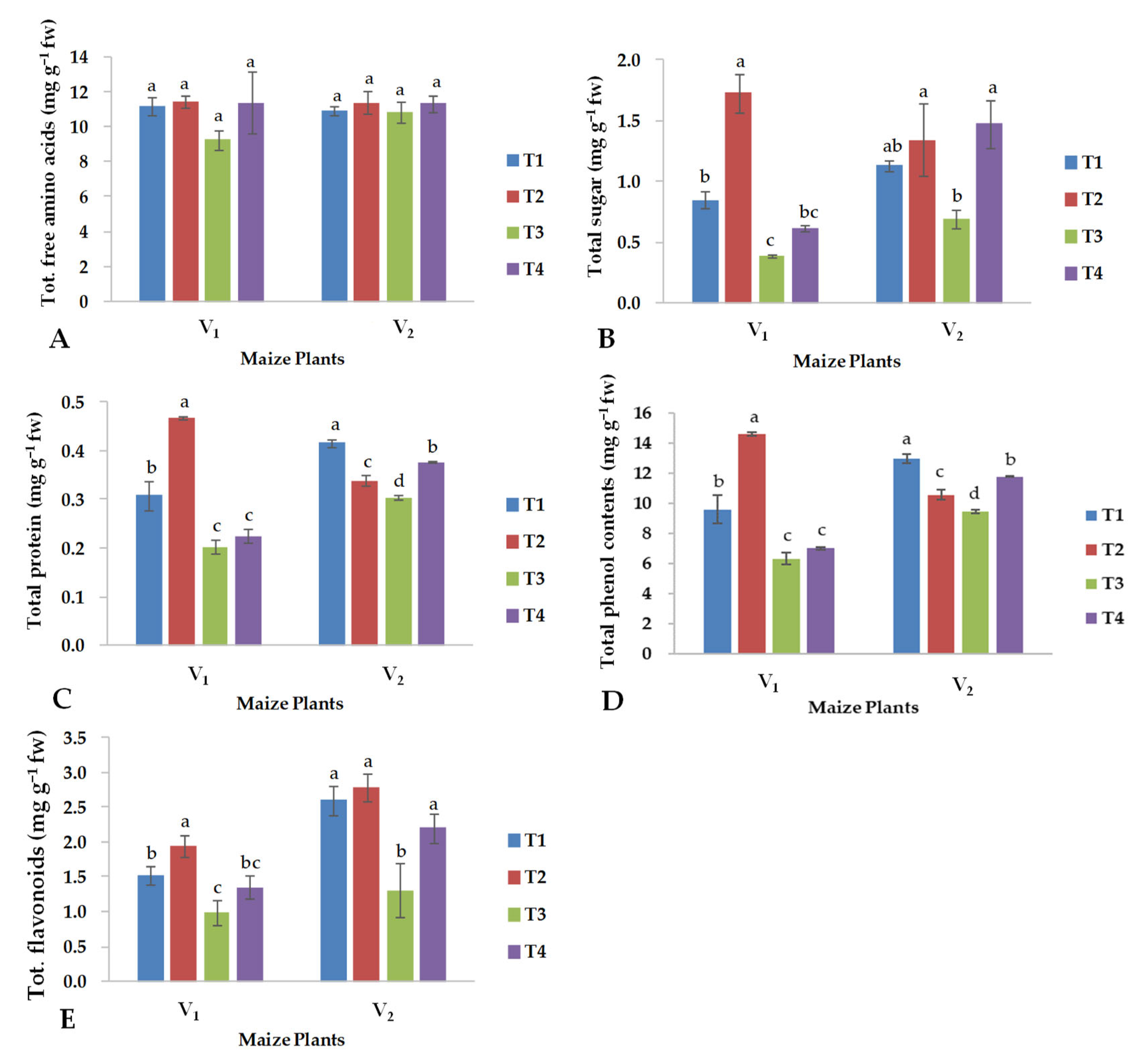
2.2. Biochemical Parameters
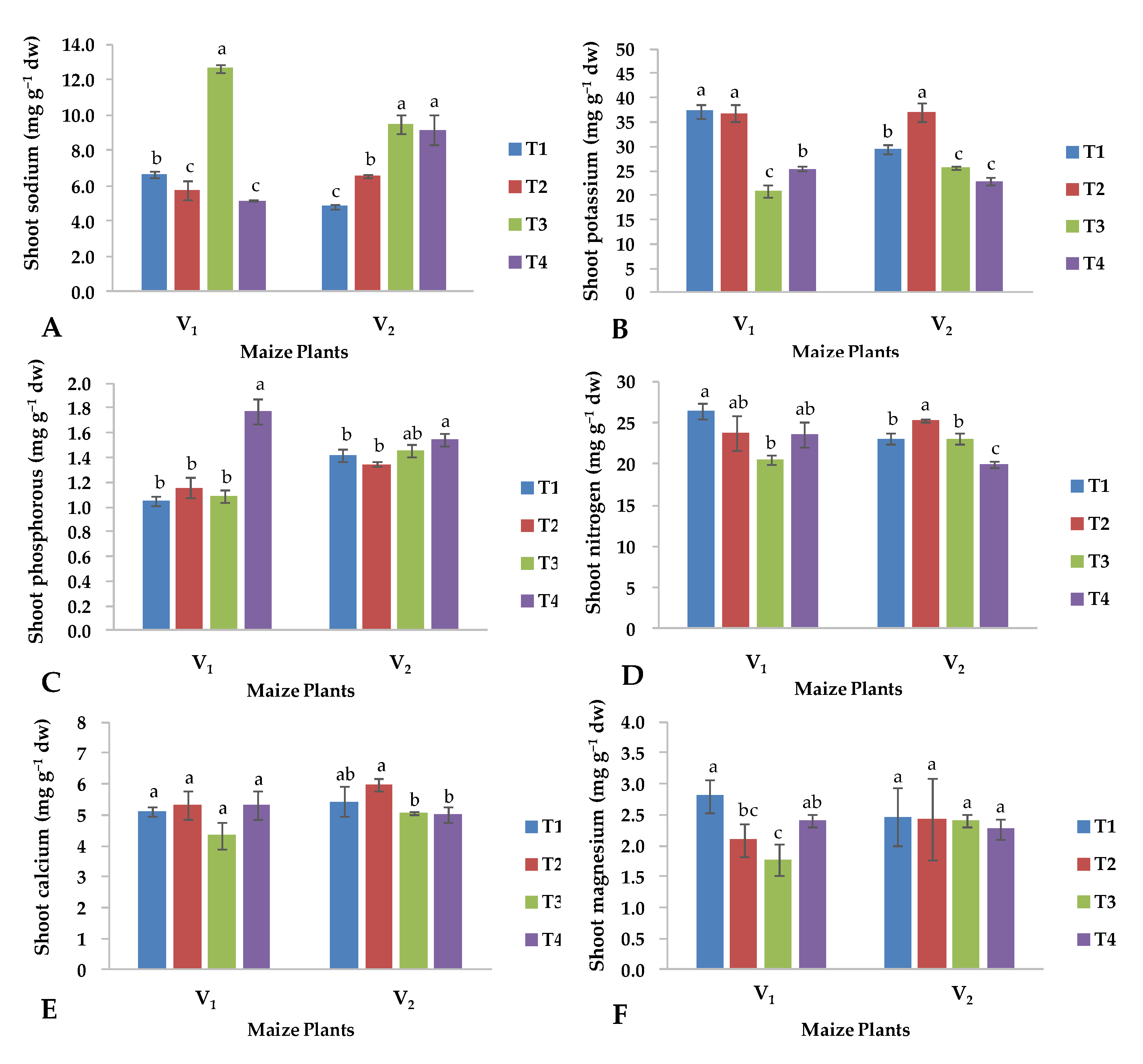
2.3. Nutrient Content in Shoot
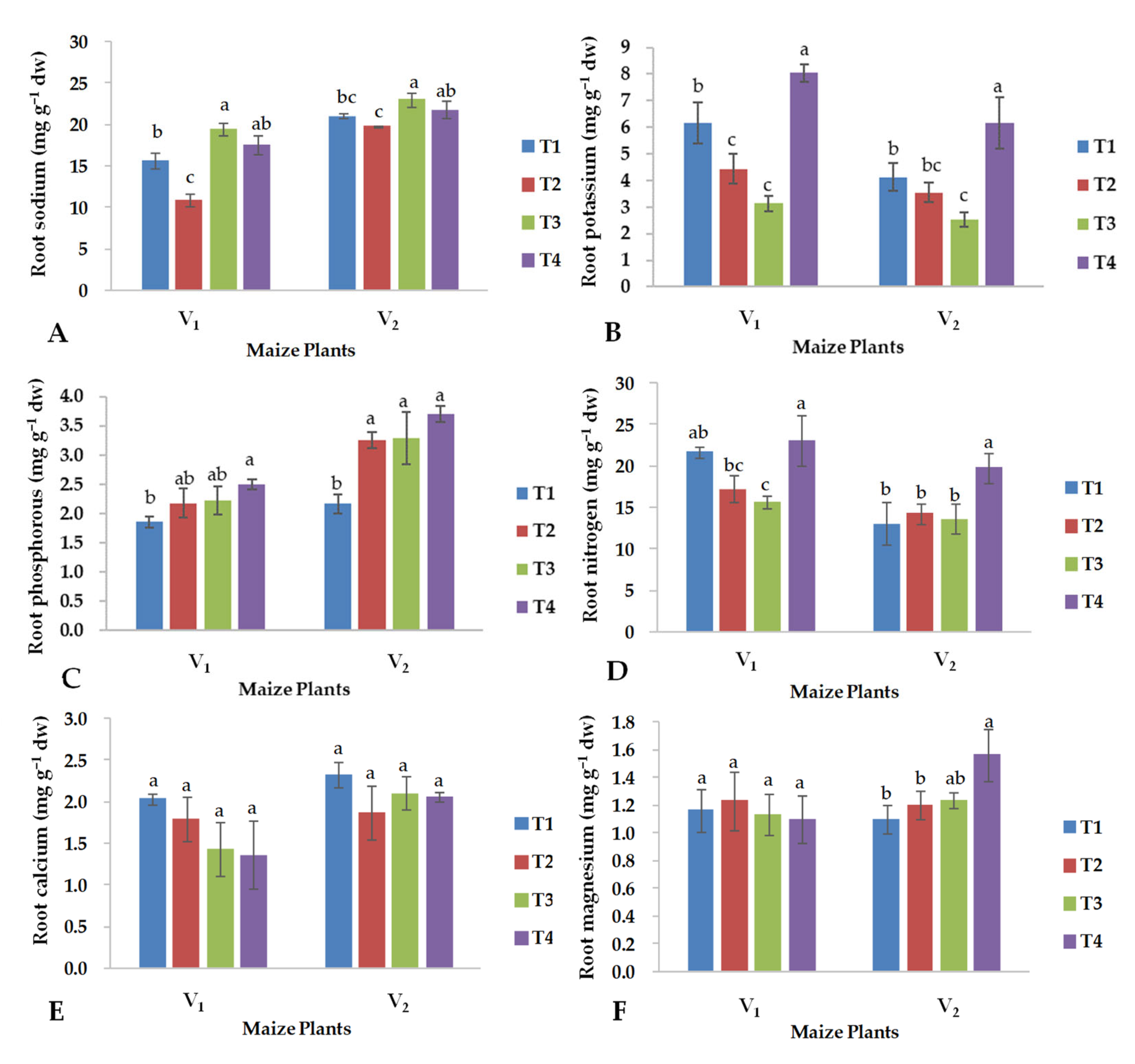
2.4. Nutrient Contents in Roots

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | DF | SL | RL | SFW | SDW | RFW | RDW | LAI | NOL | T.Chl | Chl a | Chl b | Car | TFAA | TSS | TSP | TPC | T.FAL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety | 1 | 15.360 *** | 0.260 | 0.110 | 0.032 | 0.051 | 0.014 | 597.600 *** | 35.375 ** | 0.007 | 0.002 * | 0.009 *** | 2.863 | 7.116 * | 0.423 *** | 0.020 *** | 19.429 ** | 3.600 ** |
| Salinity | 1 | 90.090 *** | 1.000 | 75.590 * | 5.587 *** | 0.111 | 0.002 ns | 4291.500 *** | 15.042 * | 0.035 ** | 0.026 *** | 0.007 *** | 0.238 | 2.537 | 1.330 *** | 0.066 *** | 64.908 *** | 3.353 ** |
| CuO-NPs | 1 | 12.610 *** | 3.154 | 0.389 | 1.540 ** | 0.002 | 0.001 | 289.820 ** | 18.375 * | 0.096 *** | 0.023 *** | 0.017 *** | 1.212 | 16.454 *** | 1.649 *** | 0.012 ** | 11.646 ** | 1.279 * |
| V × S | 1 | 3.840 * | 0.220 | 19.234 | 0.047 | 0.057 | 0.003 | 719.850 *** | 5.042 | 0.0115 | 0.004 *** | 0.0002 | 3.347 | 0.727 | 0.594 *** | 0.028 ** | 27.695 ** | 0.212 |
| V × CuONPs | 1 | 5.704 * | 0.634 | 51.202 * | 0.070 | 0.496 | 0.005 | 500.230 *** | 22.042 * | 0.007 | 0.0001 | 0.010 *** | 0.872 | 1.397 | 0.006 | 0.013 ** | 12.624 ** | 0.033 |
| S × CuO-NPs | 1 | 37.500 *** | 0.350 | 193.546 *** | 3.375 *** | 0.634 | 0.029 | 2202.630 *** | 9.375 | 0.050 ** | 0.012 *** | 0.021 *** | 2.529 | 0.002 | 0.002 | 0.0001 | 0.056 | 0.153 |
| V × S × CuO-NPs | 1 | 0.634 ns | 0.004 ns | 32.469 | 0.180 ns | 0.012 ns | 0.004 ns | 1393.090 *** | 9.375 ns | 1.064 ns | 0.0003 ns | 0.002 * | 0.768 ns | 5.177 * | 0.555 *** | 0.031 *** | 30.445 *** | 0.222 ns |
| Error | 14 | 0.706 | 0.789 | 10.372 | 0.172 | 0.252 | 0.012 | 22.230 | 2.696 | 0.005 | 0.00024 | 0.0004 | 4.450 | 0.759 | 0.014 | 0.001 | 1.178 | 0.278 |
| Source | DF | NaS | KS | PS | NS | CaS | MgS | NaR | KR | PR | NR | CaR | MgR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety | 1 | 0.179 | 11.760 * | 0.178 *** | 0.031 | 0.540 * | 0.094 | 182.216 *** | 15.058 *** | 5.006 *** | 1.076 *** | 1.175 ** | 0.082 |
| Salinity | 1 | 50.605 *** | 780.900 *** | 0.296 *** | 0.510 *** | 2.042 ** | 0.350 ns | 77.940 *** | 2.581 * | 1.929 ** | 0.129 | 0.398 * | 0.042 |
| CuO-NPs | 1 | 24.221 *** | 31.099 *** | 0.240 *** | 0.0003 | 1.402 ** | 0.020 | 31.855 ** | 19.820 *** | 1.657 ** | 0.404 ** | 0.185 | 0.082 |
| V × S | 1 | 3.293 | 34.656 *** | 0.067 ** | 0.003 | 0.042 | 0.094 | 15.730 * | 0.128 | 0.297 | 0.148 | 0.353 | 0.167 * |
| V × CuO-NPs | 1 | 44.200 *** | 0.228 | 0.227 *** | 0.006 | 0.282 | 0.004 | 6.816 | 0.519 | 0.320 | 0.072 | 0.011 | 0.060 |
| S × CuO-NPs | 1 | 34.297 *** | 10.088 * | 0.214 *** | 0.00082 | 4.168 | 0.570 * | 3.581 | 52.068 *** | 0.187 | 1.074 *** | 0.092 | 0.007 |
| V × S × CuO-NPs | 1 | 11.03 ** | 88.704 *** | 0.067 ** | 0.447 *** | 0.540 * | 0.770 * | 3.205 ns | 4.259 ** | 0.143 ns | 0.181 * | 0.020 ns | 0.042 ns |
| Error | 14 | 0.9974 | 1.618 | 0.004 | 0.011 | 0.123 | 0.119 | 1.951 | 0.3237 | 0.137 | 0.037 | 0.086 | 0.024 |
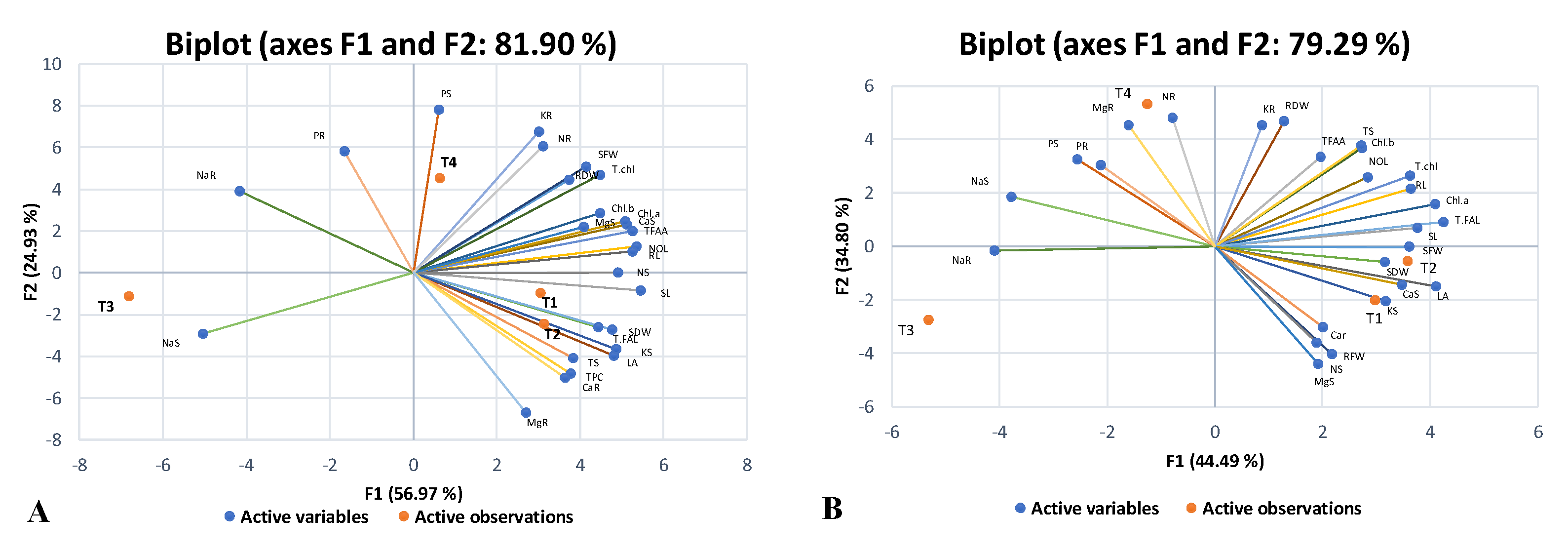
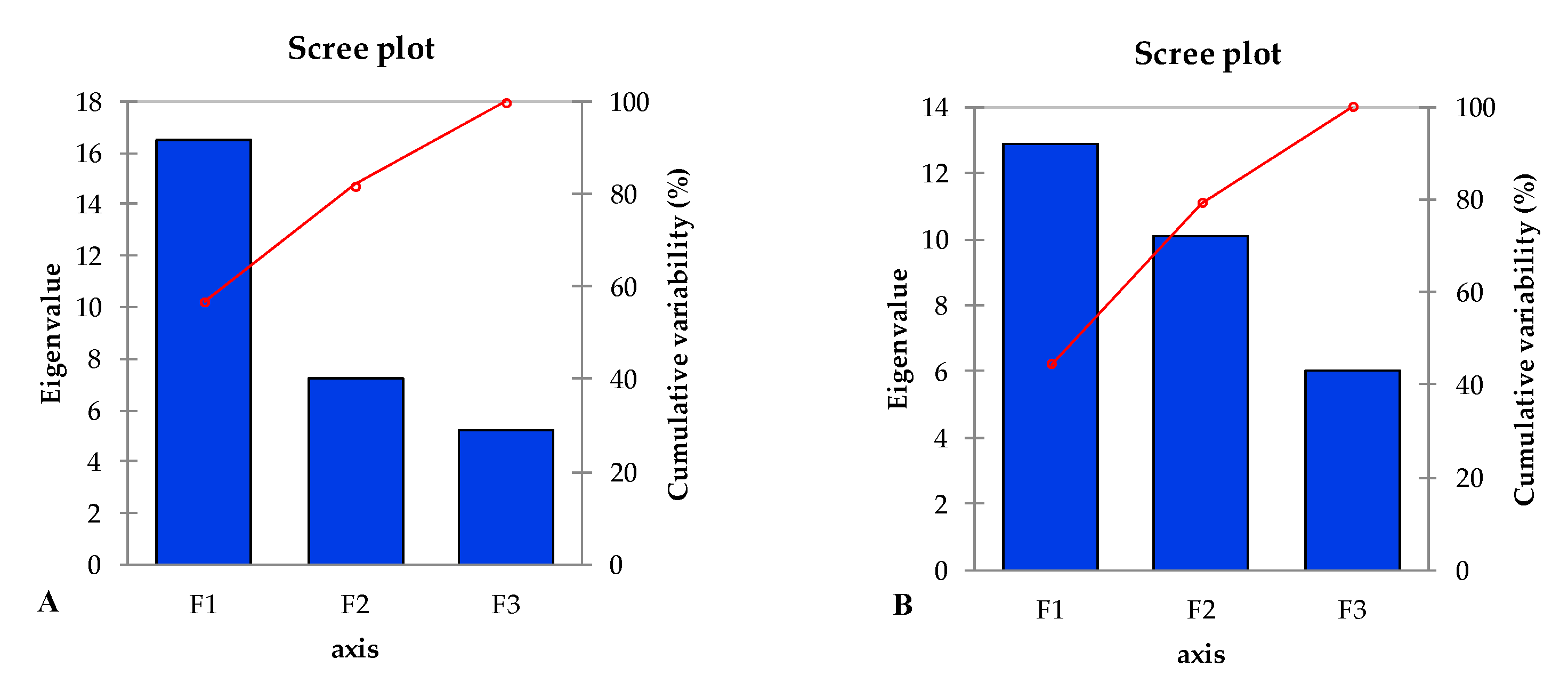
2.5. Principal Component Analysis
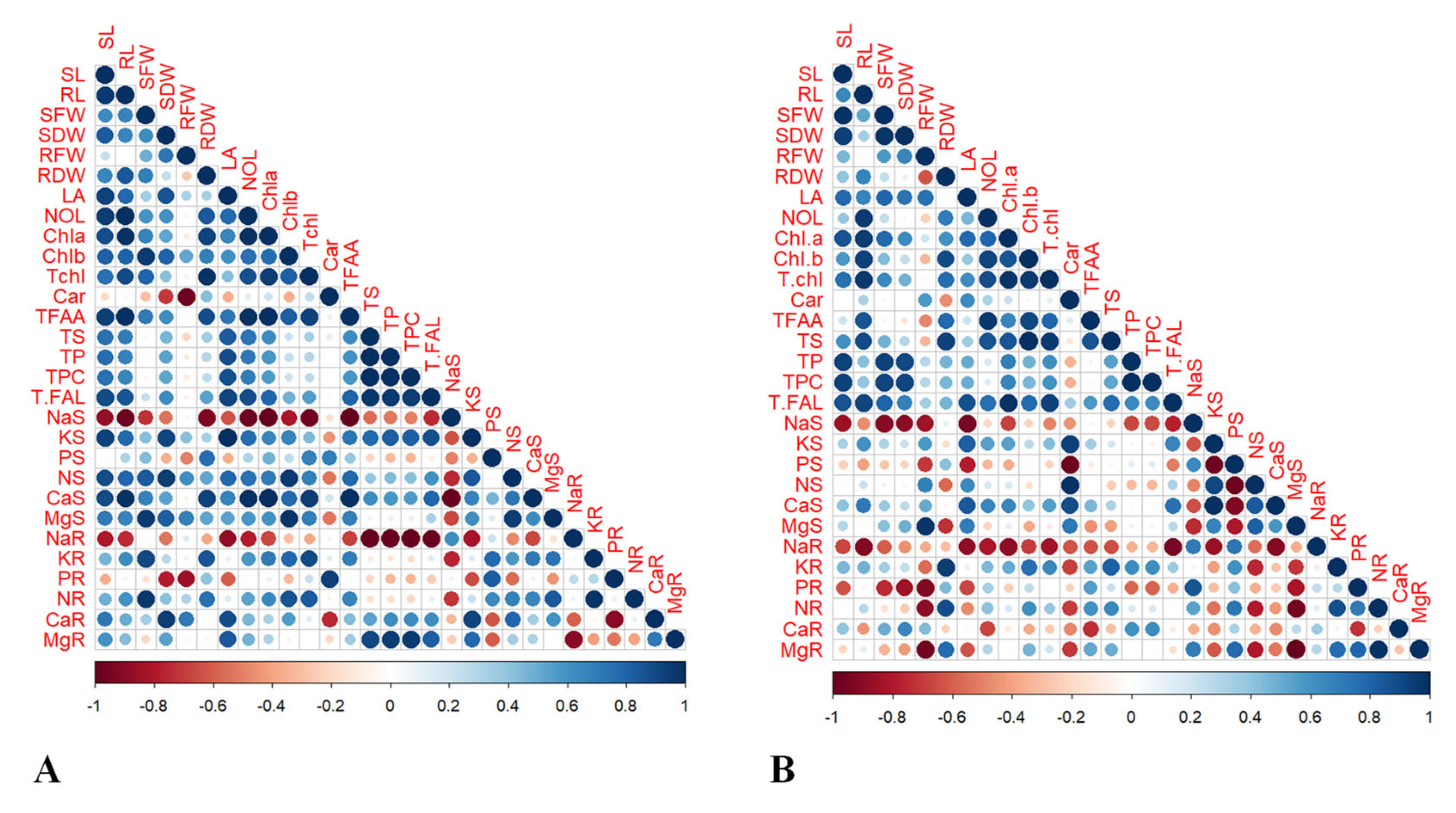
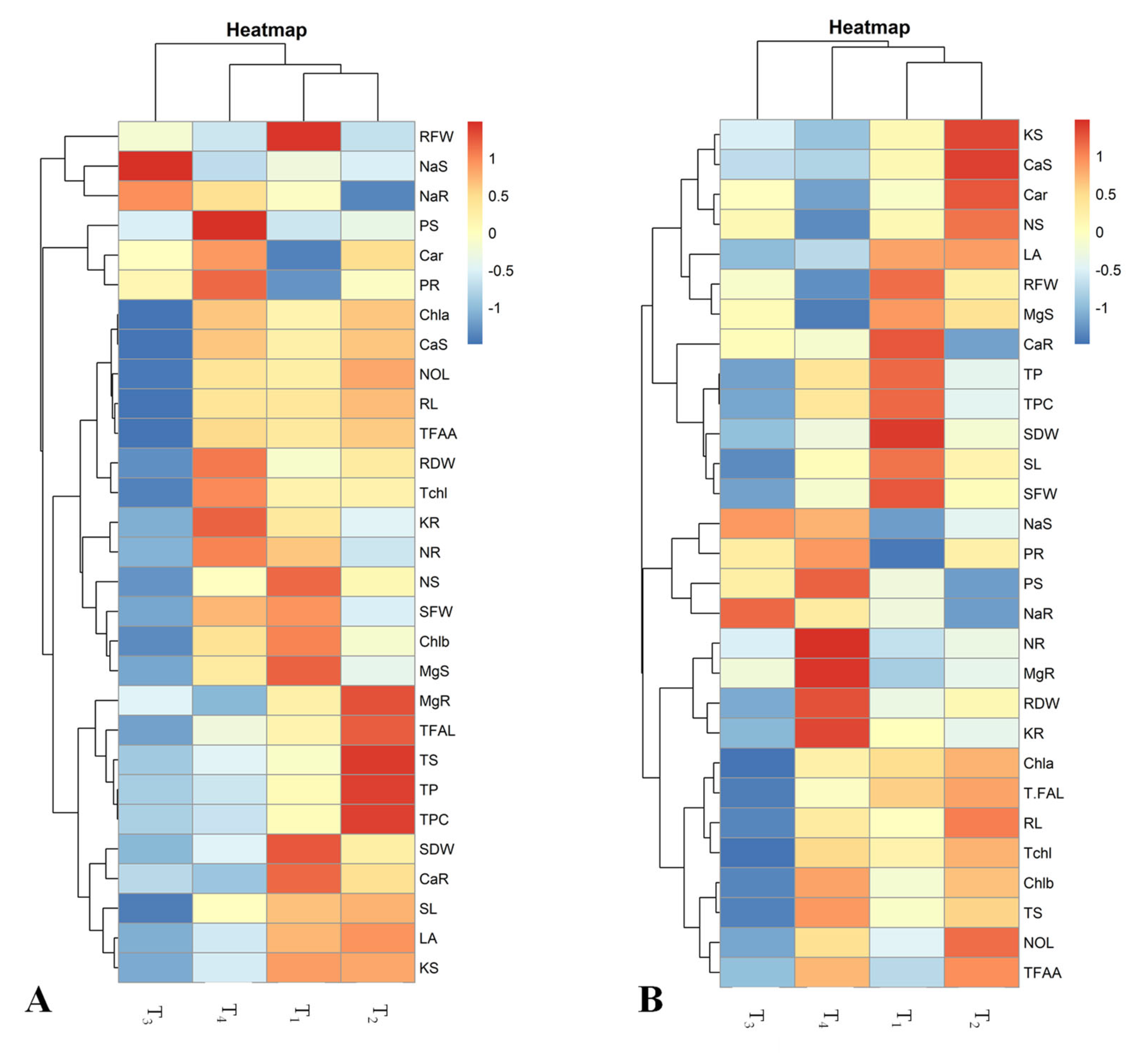
2.6. Pearson’s Correlation and Heatmap Analyses
3. Discussion
4. Materials and Methods
4.1. Experiment Layout
- (1)
- T1 (Control): irrigation with distilled water and no foliar application of CuO-NPs at two stages of development, i.e., six and nine leaves with visible leaf collars;
- (2)
- T2 (Control + CuO-NPs): irrigation with distilled water and foliar application of a 5 µM suspension of CuO-NPs at two stages of development, i.e., six and nine leaves with visible leaf collars;
- (3)
- T3 (Salinity): irrigation with a solution of NaCl with an EC value of 5 dS m−1 and no foliar application at two stages of development, i.e., six and nine leaves with visible leaf collars;
- (4)
- T4 (Salinity + CuO-NPs): irrigation with a solution of NaCl with an EC value of 5 dS m−1 and foliar application of a 5 µM suspension of CuO-NPs at two stages of development, i.e., six and nine leaves with visible leaf collars.
4.2. Growth Parameters
4.3. Biochemical Parameters
4.4. Nutrients
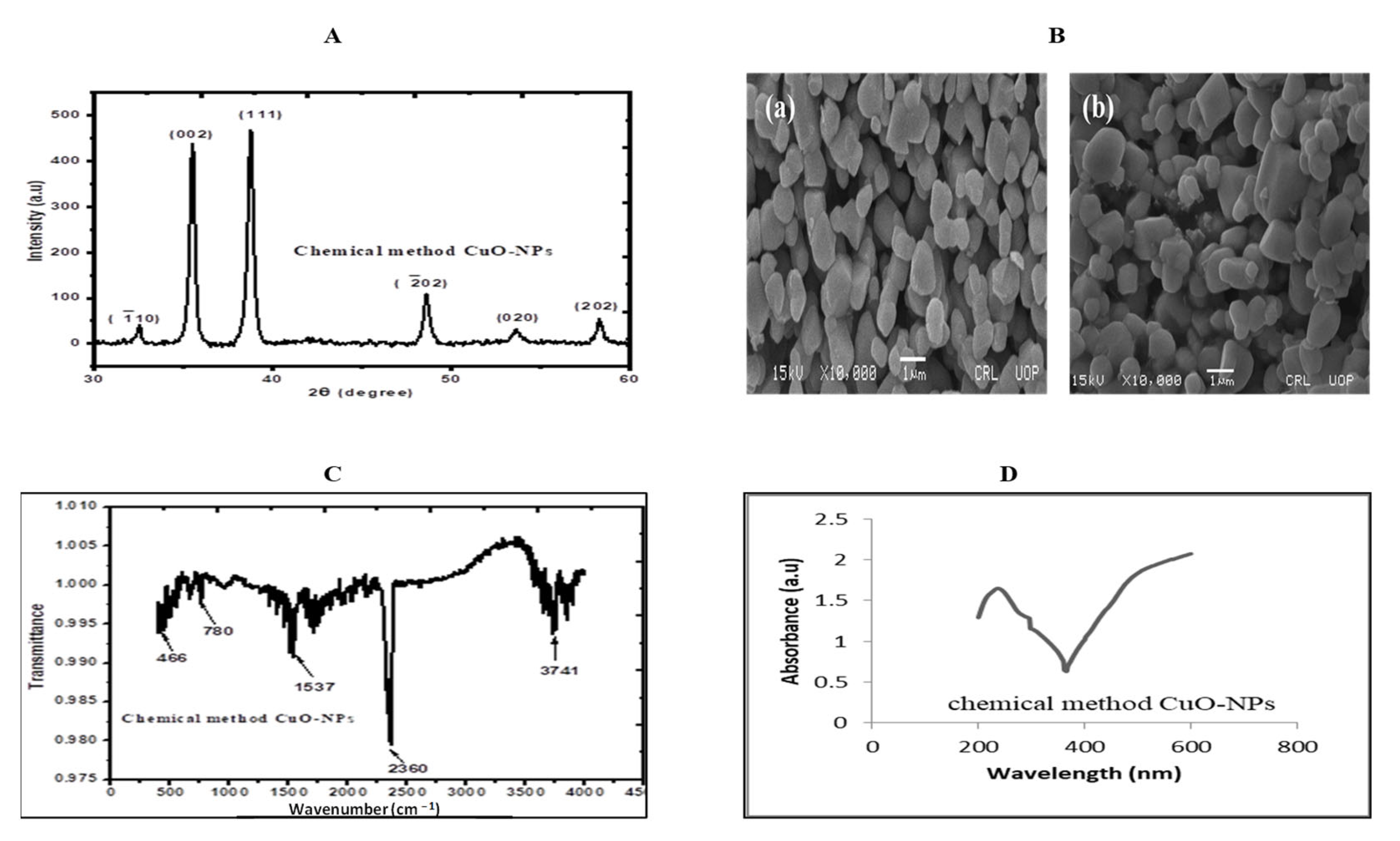
4.5. Characterization of Copper Oxide Nanoparticles
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity stress in arid and semi-arid climates: Effects and management in field crops. Clim. Change Agric. 2019, 13, 201–226. [Google Scholar]
- Faisal, M.; Faizan, M.; Tonny, S.H.; Rajput, V.D.; Minkina, T.; Alatar, A.A.; Pathirana, R. Strigolactone-Mediated Mitigation of Negative Effects of Salinity Stress in Solanum lycopersicum through Reducing the Oxidative Damage. Sustainability 2023, 15, 5805. [Google Scholar] [CrossRef]
- Negacz, K.; Malek, Ž.; de Vos, A.; Vellinga, P. Saline soils worldwide: Identifying the most promising areas for saline agriculture. J. Arid Environ. 2022, 203, 104775. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Awan, A.R.; Anwar, S.; Khaliq, B.; Malik, A.; Ozturk, M. Economic utilization of salt-affected wasteland for plant production: A case study from Pakistan. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–24. [Google Scholar]
- Ayub, M.; Ashraf, M.Y.; Kausar, A.; Saleem, S.; Anwar, S.; Altay, V.; Ozturk, M. Growth and physio-biochemical responses of maize (Zea mays L.) to drought and heat stresses. Plant Biosyst. 2021, 155, 535–542. [Google Scholar] [CrossRef]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Hussan, M.U.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar]
- Iqbal, M.N.; Rasheed, R.; Ashraf, M.Y.; Ashraf, M.A.; Hussain, I. Exogenously applied zinc and copper mitigate salinity effect in maize (Zea mays L.) by improving key physiological and biochemical attributes. Environ. Sci. Pollut. Res. 2018, 25, 23883–23896. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Ashraf, M.Y.; Khan, A.; Nawaz, F. Salicylic acid–and ascorbic acid–induced salt tolerance in mung bean (Vigna radiata (L.) Wilczek) accompanied by oxidative defense mechanisms. J. Soil Sci. Plant Nutr. 2021, 21, 2057–2071. [Google Scholar] [CrossRef]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life 2021, 11, 545. [Google Scholar] [CrossRef]
- Neshat, M.; Abbasi, A.; Hosseinzadeh, A.; Sarikhani, M.R.; Dadashi Chavan, D.; Rasoulnia, A. Plant growth promoting bacteria (PGPR) induce antioxidant tolerance against salinity stress through biochemical and physiological mechanisms. Physiol. Mol. Biol. Plants 2022, 28, 347–361. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Iqbal, N.; Ashraf, M.; Akhter, J. Modulation of physiological and biochemical metabolites in salt stressed rice by foliar application of zinc. J. Plant Nutr. 2014, 37, 447–457. [Google Scholar] [CrossRef]
- Munawar, W.; Hameed, A.; Khan, M.K.R. Differential morphophysiological and biochemical responses of cotton genotypes under various salinity stress levels during early growth stage. Front. Plant Sci. 2021, 12, 622309. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.; Kausar, A.; Zahra, N.; Hafeez, M.B.; Raza, A.; Ashraf, M.Y. Methionine-induced regulation of secondary metabolites and antioxidants in maize (Zea mays L.) subjected to salinity stress. Gesunde Pflanz. 2022, 75, 1143–1155. [Google Scholar] [CrossRef]
- Zafar, S.; Ashraf, M.Y.; Saleem, M. Shift in physiological and biochemical processes in wheat supplied with zinc and potassium under saline condition. J. Plant Nutr. 2018, 41, 19–28. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, A.; Yuan, C.; Zhao, T.; Chang, H.; Zhang, J. Transcriptome analysis of liver lipid metabolism disorders of the turbot Scophthalmus maximus in response to low salinity stress. Aquaculture 2021, 534, 736273. [Google Scholar] [CrossRef]
- Revilla, P.; Alves, M.L.; Andelković, V.; Balconi, C.; Dinis, I.; Mendes-Moreira, P.; Redaelli, R.; de Galarreta, J.I.R.; Vaz Patto, M.C.; Žilić, S.; et al. Traditional foods from maize (Zea mays L.) in Europe. Front. Nutr. 2022, 8, 683399. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; DaCosta, M.; Jiang, Y. Research advances in mechanisms of turfgrass tolerance to abiotic stresses: From physiology to molecular biology. Crit. Rev. Plant Sci. 2014, 33, 141–189. [Google Scholar] [CrossRef]
- Subbaiah, P.V.; Kumar, Y. Review on effect of inorganic and organic sources of nutrients and their integrated use on growth and yield of maize (Zea mays L.). Int. J. Conserv. Sci. 2019, 7, 896–901. [Google Scholar]
- Xu, X.; Ouyang, X.; Gu, Y.; Cheng, K.; Smith, P.; Sun, J.; Li, Y.; Pan, G. Climate change may interact with nitrogen fertilizer management leading to different ammonia loss in China’s croplands. Glob. Chang. Biol. 2021, 27, 6525–6535. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Kaur, H.; Kaur, H.; Srivastava, S. The beneficial roles of trace and ultratrace elements in plants. Plant Growth Regul. 2023, 100, 219–236. [Google Scholar] [CrossRef]
- Lodde, V.; Morandini, P.; Costa, A.; Murgia, I.; Ezquer, I. cROStalk for life: Uncovering ROS signaling in plants and animal systems, from gametogenesis to early embryonic development. Genes 2021, 12, 525. [Google Scholar] [CrossRef]
- Wang, H.; An, T.; Huang, D.; Liu, R.; Xu, B.; Zhang, S.; Deng, X.; Siddique, K.H.M.; Chen, Y. Arbuscular mycorrhizal symbioses alleviating salt stress in maize is associated with a decline in root-to-leaf gradient of Na+/K+ ratio. BMC Plant Biol. 2021, 21, 457. [Google Scholar] [CrossRef] [PubMed]
- Suganya, A.; Saravanan, A.; Manivannan, N. Role of zinc nutrition for increasing zinc availability, uptake, yield, and quality of maize (Zea mays L.) grains: An overview. Commun. Soil Sci. Plant Anal. 2020, 51, 2001–2021. [Google Scholar]
- Heinemann, B.; Künzler, P.; Eubel, H.; Braun, H.P.; Hildebrandt, T.M. Estimating the number of protein molecules in a plant cell: Protein and amino acid homeostasis during drought. Plant Physiol. 2021, 185, 385–404. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Ma, J.; Xian, J.; Peijnenburg, W.J.; Du, Y.; Tian, D.; Xiao, H.; He, Y.; Luo, L.; Deng, O.; et al. Copper accumulation and physiological markers of soybean (Glycine max) grown in agricultural soil amended with copper nanoparticles. Ecotoxicol. Environ. Saf. 2022, 229, 113088. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, A. Assessment of toxic interaction of nano zinc oxide and nano copper oxide on germination of Raphanus sativus seeds. Environ. Monit. Assess. 2019, 191, 703. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Choudhary, A.; Kaur, H.; Mehta, S.; Husen, A. Metal-based nanoparticles, sensors, and their multifaceted application in food packaging. J. Nanobiotechnol. 2021, 19, 256. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture—Recent developments and future perspectives. J. Nanobiotechnol. 2022, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Seregina, T.; Chernikova, O.; Mazhaysky, Y.; Ampleeva, L. Features of the influence of copper nanoparticles and copper oxide on the formation of barley crop. Agron. Res. 2020, 18, 1010–1017. [Google Scholar]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.; Sharma, N.; Pankaj Sahu, P.; Prasad, M. Chromatin-based epigenetic regulation of plant abiotic stress response. Curr. Genom. 2016, 17, 490–498. [Google Scholar] [CrossRef]
- Trotta, F.; Mele, A. Nanomaterials: Classification and Properties. In Nanosponges: Synthesis and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 1–26. [Google Scholar]
- Shani, M.Y.; Ahmed, S.R.; Ashraf, M.Y.; Khan, Z.; Cocozza, C.; De Mastro, F.; Gul, N.; Pervaiz, S.; Abbas, S.; Nawaz, H.; et al. Nano-Biochar Suspension Mediated Alterations in Yield and Juice Quality of Kinnow (Citrus reticulata L.). Horticulturae 2023, 9, 521. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, C.; Wang, H.; Han, Y.; Tai, F.; Wu, J.; Li, H.; He, R. Biphasic impacts of graphite-derived engineering carbon-based nanomaterials on plant performance: Effectiveness vs. nanotoxicity. Adv. Agrochem. 2023, 2, 113–126. [Google Scholar] [CrossRef]
- Yadav, M.; George, N.; Dwibedi, V. Emergence of toxic trace elements in plant environments: Insights into potential of silica nanoparticles for mitigation of metal toxicity in plants. Environ. Pollut. 2023, 333, 122112. [Google Scholar] [CrossRef]
- Khalid, M.F.; Iqbal Khan, R.; Jawaid, M.Z.; Shafqat, W.; Hussain, S.; Ahmed, T.; Rizwan, M.; Ercisli, S.; Pop, O.L.; Marc, A.R. Nanoparticles: The plant saviour under abiotic stresses. Nanomaterials 2022, 12, 3915. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.H.; Sabir, I.A.; Rehman, A.; Hameed, M.K.; Albashar, G.; Manzoor, M.A.; Shakoor, A. Co-application of copper oxide nanoparticles and Trichoderma harzianum with physiological, enzymatic and ultrastructural responses for the mitigation of salt stress. Chemosphere 2023, 336, 139230. [Google Scholar] [CrossRef]
- Abbasi, G.H.; Akhtar, J.; Ahmad, R.; Jamil, M.; Anwar-ul-Haq, M.; Ali, S.; Ijaz, M. Potassium application mitigates salt stress differentially at different growth stages intolerant and sensitive maize hybrids. Plant Growth Regul. 2015, 76, 111–125. [Google Scholar] [CrossRef]
- Khalil, S.E. Review: Roles of Some Abiotic Stresses on Plant Growth Characters. J. Mod. Agric. Biotechnol. 2022, 1, 9. [Google Scholar] [CrossRef]
- Raza, A.; Salehi, H.; Rahman, M.A.; Zahid, Z.; Madadkar Haghjou, M.; Najafi-Kakavand, S.; Charagh, S.; Osman, H.S.; Albaqami, M.; Zhuang, Y.; et al. Plant hormones and neurotransmitter interactions mediate antioxidant defenses under induced oxidative stress in plants. Front. Plant Sci. 2022, 13, 961872. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Tripathi, D.K.; Chauhan, D.K.; Sharma, S.; Sahi, S. Potential applications and avenues of nanotechnology in sustainable agriculture. In Nanomaterials in Plants, Algae, and Microorganisms; Academic Press: Cambridge, MA, USA, 2018; pp. 473–500. [Google Scholar]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef]
- Nadal, M.; Carriquí, M.; Flexas, J. Chapter 3 Mesophyll conductance to CO2 diffusion in a climate change scenario: Effects of elevated CO2, temperature and water stress. In Photosynthesis, Respiration, and Climate Change; Springer International Publishing: Cham, Switzerland, 2021; pp. 49–78. [Google Scholar]
- Singh, P.; Kumar, V.; Sharma, J.; Saini, S.; Sharma, P.; Kumar, S.; Sinhmar, Y.; Kumar, D.; Sharma, A. Silicon Supplementation Alleviates the Salinity Stress in Wheat Plants by Enhancing the Plant Water Status, Photosynthetic Pigments, Proline Content and Antioxidant Enzyme Activities. Plants 2022, 11, 2525. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Li, G.; Xie, D.; Farag, R.; Hasan, M.M.; Alabdallah, N.M.; Al-Harbi, N.A.; Al-Qahtani, S.M.; Zeeshan, M.; Nasar, J.; et al. Melatonin Mitigates Salt-Induced Growth Inhibition through the Regulation of Carbohydrate and Nitrogen Metabolism in Tomato Seedlings. J. Soil Sci. Plant Nutr. 2023, 23, 4290–4308. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Diao, P.; Kong, L.; Yu, R.; Zhang, M.; Zuo, T.; Niu, Y.; Fan, Y.; Wuriyanghan, H. Ethylene enhances seed germination and seedling growth under salinity by reducing oxidative stress and promoting chlorophyll content via ETR2 pathway. Front. Plant Sci. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Ashraful Alam, M.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Consequences and mitigation strategies of abiotic stresses in wheat (Triticum aestivum L.) under the changing climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Karumannil, S.; Khan, T.A.; Kappachery, S.; Gururani, M.A. Impact of exogenous melatonin application on photosynthetic machinery under abiotic stress conditions. Plants 2023, 12, 2948. [Google Scholar] [CrossRef] [PubMed]
- Al-Hakimi, A.B.; Hamada, A.M. Ascorbic acid, thiamine or salicylic acid induced changes in some physiological parameters in wheat grown under copper stress. Plant Soil Environ. 2011, 47, 92–108. [Google Scholar] [CrossRef]
- Zengin, F.K.; Sevda, K. Effects of copper on chlorophyll, proline, protein and abscisic acid level of sunflower (Helianthus annuus L.) seedlings. J. Environ. Biol. 2007, 28, 561. [Google Scholar]
- Zhang, Y.; Sun, X.; Aphalo, P.J.; Zhang, Y.; Cheng, R.; Li, T. Ultraviolet-A1 radiation induced a more favorable light-intercepting leaf-area display than blue light and promoted plant growth. Plant Cell Environ. 2023, 47, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Gill, S.; Ramzan, M.; Ahmad, M.Z.; Danish, S.; Huang, P.; Obaid, S.A.; Alharbi, S.A. Uncovering the impact of AM fungi on wheat nutrient uptake, ion homeostasis, oxidative stress, and antioxidant defense under salinity stress. Sci. Rep. 2023, 13, 8249. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.M.; Pradhan, N.; Subba, R.; Saha, P.; Roy, S. Sugar-terminated carbon-nanodots stimulate osmolyte accumulation and ROS detoxification for the alleviation of salinity stress in Vigna radiata. Sci. Rep. 2022, 12, 17567. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, N.; Geetha, N.; Ghorbanpour, M.; Venkatachalam, P. Role of Engineered Zinc and Copper Oxide Nanoparticles in Promoting Plant Growth and Yield: Present Status and Future Prospects. In Advances in Phytonanotechnology: From Synthesis to Application; Elsevier: Amsterdam, The Netherlands, 2019; pp. 183–201. [Google Scholar]
- Hassanein, A.; Esmail, N.; Hashem, H. Sodium nitroprusside mitigates the inhibitory effect of salt and heavy metal stress on lupine yield and downregulates antioxidant enzyme activities. Acta Agrobot. 2020, 73, 7336. [Google Scholar] [CrossRef]
- Aldesuquy, H.; Baka, Z.; Mickky, B. Kinetin and spermine mediated induction of salt tolerance in wheat plants: Leaf area, photosynthesis and chloroplast ultrastructure of flag leaf at ear emergence. Egypt. J. Basic Appl. Sci. 2014, 1, 77–87. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Davies, B.H. Carotenoids. In Chemistry and Biochemistry of Plant Pigments; Goodwin, T., Ed.; Academic Press: London, UK, 1976; Volume 2, pp. 38–165. [Google Scholar]
- Riazi, A.; Matsuda, K.; Arslan, A. Water-stress induced changes in concentrations of proline and other solutes in growing regions of young barley leaves. J. Exp. Bot. 1985, 36, 1716–1725. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 191, 265–275. [Google Scholar] [CrossRef]
- Hamilton, P.B.; Van Slyke, D.D. The gasometric determination of free amino acids in blood filtrates by the ninhydrin-carbon dioxide method. J. Biol. Chem. 1943, 150, 231–250. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic–phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Sofowora, A. Medicinal Plants and Traditional Medicine in Africa; Spectrum Books Ltd.: Ibadan, Nigeria, 1993; Volume 289. [Google Scholar]
- Harborne, I.B. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis, 2nd ed.; Chapman and Hall: New York, NY, USA, 1973; pp. 88–185. [Google Scholar]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall: Hoboken, NJ, USA, 1958; p. 30. [Google Scholar]
- Bremner, J.M.; Edwards, A.P. Determination and isotope-ratio analysis of different forms of nitrogen in soils: I. Apparatus and procedure for distillation and determination of ammonium. Soil Sci. Soc. Am. J. 1965, 29, 504–507. [Google Scholar] [CrossRef]
- Haider, H.I.; Zafar, I.; Ain, Q.U.; Noreen, A.; Nazir, A.; Javed, R.; Sehgal, S.A.; Khan, A.A.; Rahman, M.M.; Rashid, S.; et al. Synthesis and characterization of copper oxide nanoparticles: Its influence on corn (Z. mays) and wheat (Triticum aestivum) plants by inoculation of Bacillus subtilis. Environ. Sci. Pollut. Res. Int. 2022, 30, 37370–37385. [Google Scholar] [CrossRef] [PubMed]










| Soil Characteristics | Values |
|---|---|
| Soil Texture | Loamy |
| EC (dS m−1) | 0.5 |
| pH | 7.8 |
| Organic matter (%) | 0.4 |
| Available Phosphorus (ppm) | 2.4 |
| Extractable Potassium (ppm) | 45.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafiq, H.; Shani, M.Y.; Ashraf, M.Y.; De Mastro, F.; Cocozza, C.; Abbas, S.; Ali, N.; Zaib-un-Nisa; Tahir, A.; Iqbal, M.; et al. Copper Oxide Nanoparticles Induced Growth and Physio-Biochemical Changes in Maize (Zea mays L.) in Saline Soil. Plants 2024, 13, 1080. https://doi.org/10.3390/plants13081080
Shafiq H, Shani MY, Ashraf MY, De Mastro F, Cocozza C, Abbas S, Ali N, Zaib-un-Nisa, Tahir A, Iqbal M, et al. Copper Oxide Nanoparticles Induced Growth and Physio-Biochemical Changes in Maize (Zea mays L.) in Saline Soil. Plants. 2024; 13(8):1080. https://doi.org/10.3390/plants13081080
Chicago/Turabian StyleShafiq, Hina, Muhammad Yousaf Shani, Muhammad Yasin Ashraf, Francesco De Mastro, Claudio Cocozza, Shahid Abbas, Naila Ali, Zaib-un-Nisa, Aqsa Tahir, Muhammad Iqbal, and et al. 2024. "Copper Oxide Nanoparticles Induced Growth and Physio-Biochemical Changes in Maize (Zea mays L.) in Saline Soil" Plants 13, no. 8: 1080. https://doi.org/10.3390/plants13081080