
Transcriptome Analysis Reveals Key Genes Involved in Trichome Formation in Pepper (Capsicum annuum L.)

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Observations on the Epidermal Trichomes of Peppers
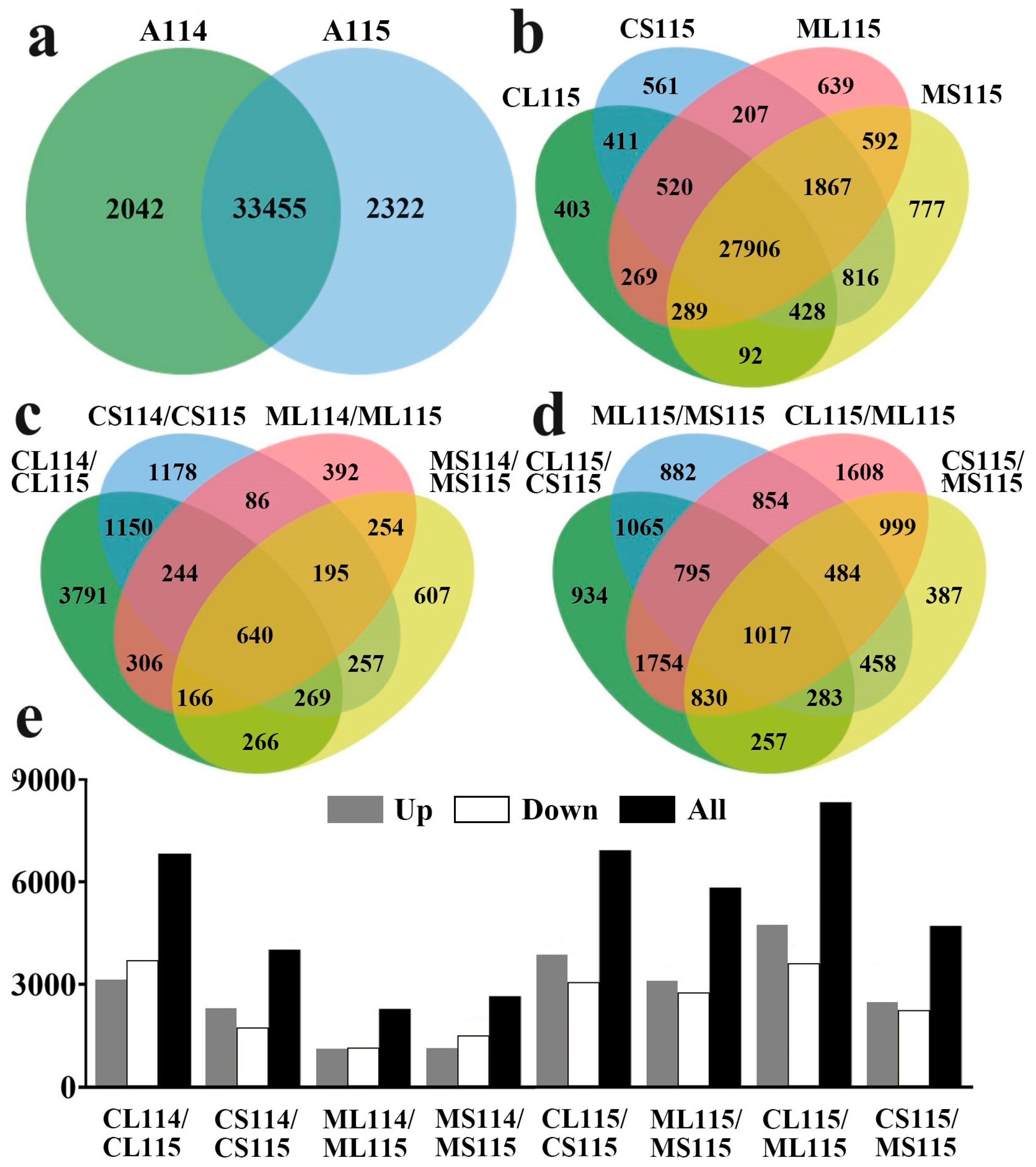
2.2. Illumina-Based Transcriptome Analysis and Identification of DEGs
2.3. GO Enrichment Analysis of DEGs
2.4. KEGG Pathway Analysis of DEGs
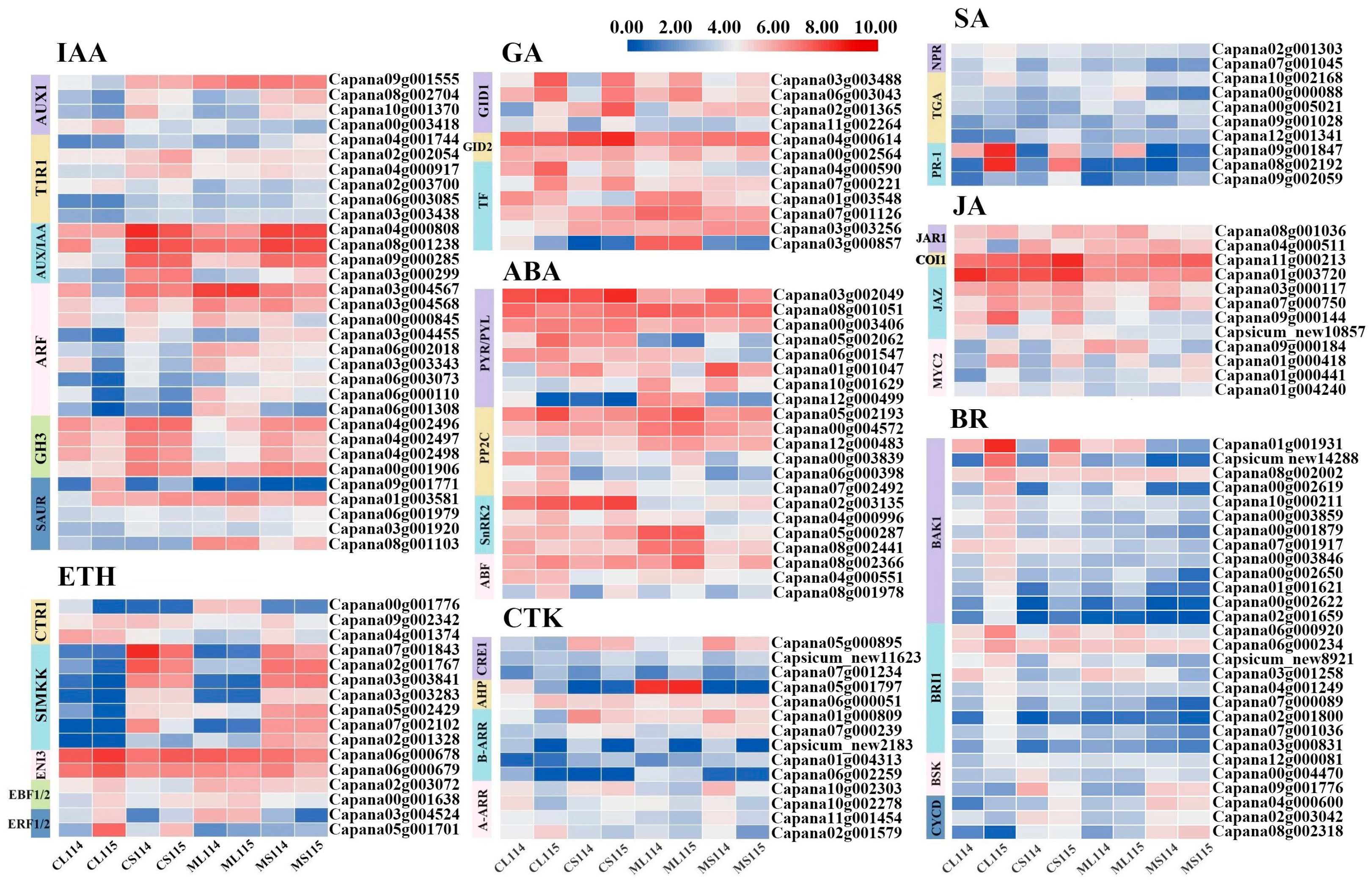
2.5. Analysis of DEGs Associated with Plant Hormone Signal Transduction Pathways
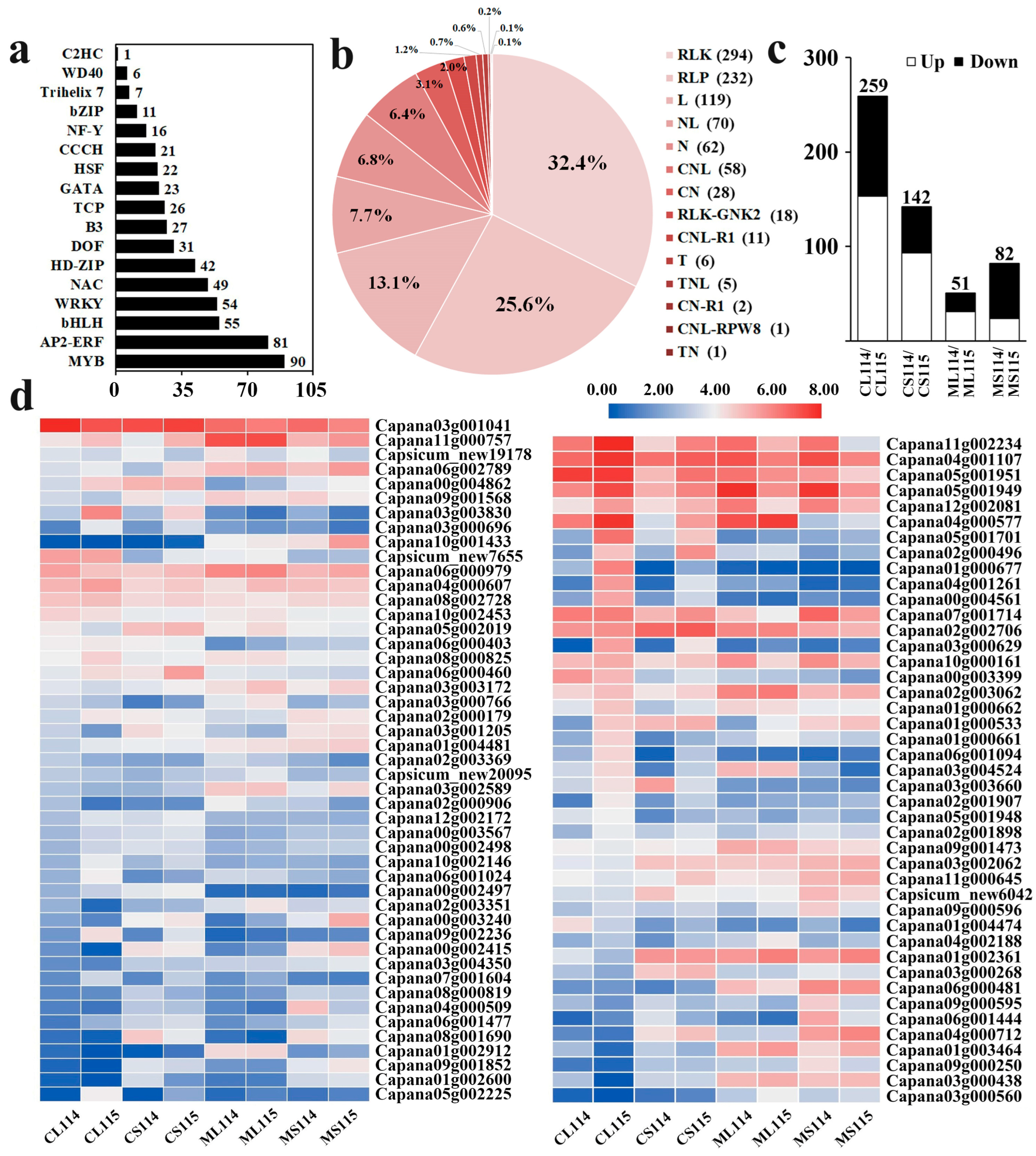
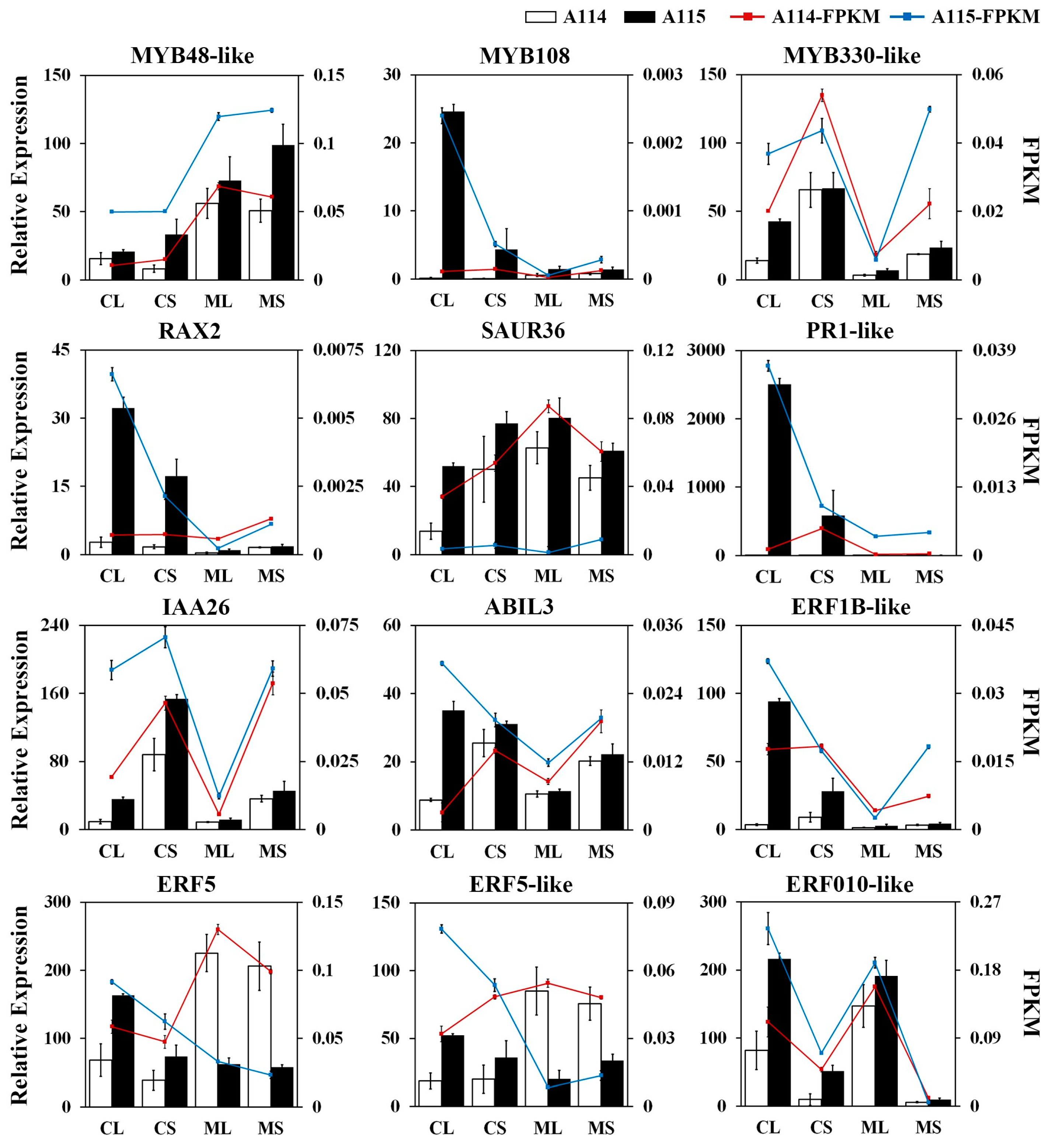
2.6. Analysis of Transcription Factors Associated with Trichome Development
2.7. Analysis of DEGs Associated with Plant Resistance
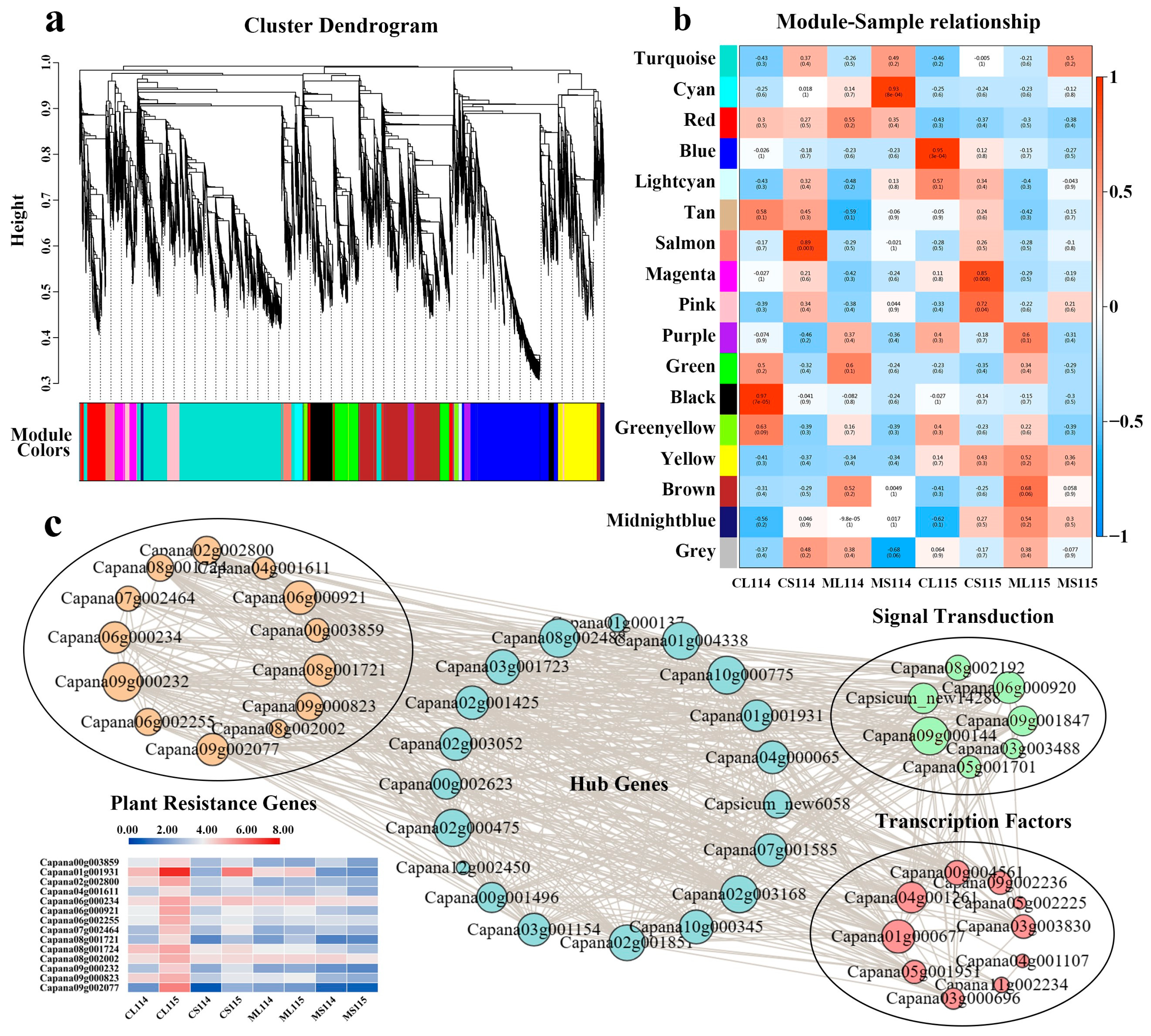
2.8. Analysis of Co-Expression Associated with Trichome Development
2.9. Analysis of Co-Expression Associated with Trichome Development
3. Discussion
4. Materials and Methods
4.1. Plant Material and Phenotypic Observations
4.2. RNA Extraction, Library Construction, and Sequencing
4.3. RNA-Seq and Differential Expression Analysis
4.4. GO and KEGG Pathway Analysis
4.5. Weighted Gene Co-Expression Network Analysis (WGCNA)
4.6. Quantitative RT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Wang, X.; Jin, Z.; Müller, C.; Pugh, T.A.M.; Chen, A.; Wang, T.; Huang, L.; Zhang, Y.; Li, L.X.Z.; et al. Occurrence of Crop Pests and Diseases Has Largely Increased in China since 1970. Nat. Food 2022, 3, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.F.; Gil, F.; Lacasaña, M.; Rodríguez-Barranco, M.; Tsatsakis, A.M.; Requena, M.; Parrón, T.; Alarcón, R. Pesticide Exposure and Genetic Variation in Xenobiotic-Metabolizing Enzymes Interact to Induce Biochemical Liver Damage. Food Chem. Toxicol. 2013, 61, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef]
- Kariyat, R.R.; Balogh, C.M.; Moraski, R.P.; De Moraes, C.M.; Mescher, M.C.; Stephenson, A.G. Constitutive and Herbivore-Induced Structural Defenses Are Compromised by Inbreeding in Solanum carolinense (Solanaceae). Am. J. Bot. 2013, 100, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tosens, T.; Harley, P.C.; Jiang, Y.; Kanagendran, A.; Grosberg, M.; Jaamets, K.; Niinemets, Ü. Glandular Trichomes as a Barrier against Atmospheric Oxidative Stress: Relationships with Ozone Uptake, Leaf Damage, and Emission of LOX Products across a Diverse Set of Species. Plant Cell Environ. 2018, 41, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Kariyat, R.R.; Hardison, S.B.; Ryan, A.B.; Stephenson, A.G.; De Moraes, C.M.; Mescher, M.C. Leaf Trichomes Affect Caterpillar Feeding in an Instar-Specific Manner. Commun. Integr. Biol. 2018, 11, 1–6. [Google Scholar] [CrossRef]
- Firdaus, S.; Van Heusden, A.; Harpenas, A.; Supena, E.D.J.; Visser, R.G.F.; Vosman, B. Identification of Silverleaf Whitefly Resistance in Pepper. Plant Breed. 2011, 130, 708–714. [Google Scholar] [CrossRef]
- Kim, H.J.; Han, J.-H.; Kim, S.; Lee, H.R.; Shin, J.-S.; Kim, J.-H.; Cho, J.; Kim, Y.H.; Lee, H.J.; Kim, B.-D.; et al. Trichome Density of Main Stem Is Tightly Linked to PepMoV Resistance in Chili Pepper (Capsicum annuum L.). Theor. Appl. Genet. 2011, 122, 1051–1058. [Google Scholar] [CrossRef]
- Lu, F.-H.; Kwon, S.-W.; Yoon, M.-Y.; Kim, K.-T.; Cho, M.-C.; Yoon, M.-K.; Park, Y.-J. SNP Marker Integration and QTL Analysis of 12 Agronomic and Morphological Traits in F8 RILs of Pepper (Capsicum annuum L.). Mol. Cells 2012, 34, 25–34. [Google Scholar] [CrossRef]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and Review of Trichomes in Plants. BMC Plant Biol. 2021, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Morohashi, K.; Hatlestad, G.; Grotewold, E.; Lloyd, A. The TTG1-bHLH-MYB Complex Controls Trichome Cell Fate and Patterning through Direct Targeting of Regulatory Loci. Development 2008, 135, 1991–1999. [Google Scholar] [CrossRef] [PubMed]
- Doroshkov, A.V.; Konstantinov, D.K.; Afonnikov, D.A.; Gunbin, K.V. The Evolution of Gene Regulatory Networks Controlling Arabidopsis thaliana L. Trichome Development. BMC Plant Biol. 2019, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.; Smith, K.; Tierney, I.; MacKenzie, K.; Hackett, C.A. Mapping Gene H Controlling Cane Pubescence in Raspberry and Its Association with Resistance to Cane Botrytis and Spur Blight, Rust and Cane Spot. Theor. Appl. Genet. 2006, 112, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Khosla, A.; Paper, J.M.; Boehler, A.P.; Bradley, A.M.; Neumann, T.R.; Schrick, K. HD-Zip Proteins GL2 and HDG11 Have Redundant Functions in Arabidopsis Trichomes, and GL2 Activates a Positive Feedback Loop via MYB23. Plant Cell 2014, 26, 2184–2200. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Cai, W.-J.; Wang, S.; Shan, C.-M.; Wang, L.-J.; Chen, X.-Y. Temporal Control of Trichome Distribution by MicroRNA156-Targeted SPL Genes in Arabidopsis Thaliana. Plant Cell 2010, 22, 2322–2335. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Xu, Z.; Li, M.; Yang, M.; Qin, H.; Yang, J.; Wu, S. Spatiotemporal Cytoskeleton Organizations Determine Morphogenesis of Multicellular Trichomes in Tomato. PLoS Genet. 2019, 15, e1008438. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, H.; Zhang, J.; Luo, Z.; Gong, P.; Zhang, C.; Li, J.; Wang, T.; Zhang, Y.; Lu, Y.; et al. A Regulatory Gene Induces Trichome Formation and Embryo Lethality in Tomato. Proc. Natl. Acad. Sci. USA 2011, 108, 11836–11841. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Gao, Y.; Xiong, C.; Yu, G.; Chang, J.; Yang, Q.; Yang, C.; Ye, Z. The Tomato B-Type Cyclin Gene, SlCycB2, Plays Key Roles in Reproductive Organ Development, Trichome Initiation, Terpenoids Biosynthesis and Prodenia Litura Defense. Plant Sci. 2017, 262, 103–114. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; Lian, H.; Chen, H. Molecular Cloning and Characterization of Anthocyanin Biosynthesis Genes in Eggplant (Solanum melongena L.). Acta Physiol. Plant. 2016, 38, 163. [Google Scholar] [CrossRef]
- Deng, W.; Yang, Y.; Ren, Z.; Audran-Delalande, C.; Mila, I.; Wang, X.; Song, H.; Hu, Y.; Bouzayen, M.; Li, Z. The Tomato SlIAA15 Is Involved in Trichome Formation and Axillary Shoot Development. New Phytol. 2012, 194, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yan, F.; Tang, Y.; Yuan, Y.; Deng, W.; Li, Z. Auxin Response Gene SlARF3 Plays Multiple Roles in Tomato Development and Is Involved in the Formation of Epidermal Cells and Trichomes. Plant Cell Physiol. 2015, 56, 2110–2124. [Google Scholar] [CrossRef]
- Hua, B.; Chang, J.; Xu, Z.; Han, X.; Xu, M.; Yang, M.; Yang, C.; Ye, Z.; Wu, S. HOMEODOMAIN PROTEIN8 Mediates Jasmonate-Triggered Trichome Elongation in Tomato. New Phytol. 2021, 230, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luo, Y.; Zhang, W.; Jian, W.; Zhang, L.; Gao, X.; Hu, X.; Yuan, Y.; Wu, M.; Xu, X.; et al. A SlMYB75-Centred Transcriptional Cascade Regulates Trichome Formation and Sesquiterpene Accumulation in Tomato. J. Exp. Bot. 2021, 72, 3806–3820. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Seo, E.; Kim, J.; Cheong, H.-J.; Kang, B.-C.; Choi, D. Morphological Classification of Trichomes Associated with Possible Biotic Stress Resistance in the Genus Capsicum. Plant Pathol. J. 2012, 28, 107–113. [Google Scholar] [CrossRef]
- Chunthawodtiporn, J.; Hill, T.; Stoffel, K.; Deynze, A. Quantitative Trait Loci Controlling Fruit Size and Other Horticultural Traits in Bell Pepper (Capsicum annuum). Plant Genome 2018, 11, 160125. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Liu, M.; Liu, J.; Liu, S.; Cheng, Q.; Shen, H. Hairiness Gene Regulated Multicellular, Non-Glandular Trichome Formation in Pepper Species. Front. Plant Sci. 2021, 12, 784755. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, N.; Niran, J.; Wang, F.; Yin, Y.; Yu, C.; Jiao, C.; Yang, C.; Yao, M. Transcriptome Profiling of Capsicum Annuum Using Illumina- and PacBio SMRT-Based RNA-Seq for in-Depth Understanding of Genes Involved in Trichome Formation. Sci. Rep. 2021, 11, 10164. [Google Scholar] [CrossRef]
- Maes, L.; Goossens, A. Hormone-Mediated Promotion of Trichome Initiation in Plants Is Conserved but Utilizes Species- and Trichome-Specific Regulatory Mechanisms. Plant Signal. Behav. 2010, 5, 205–207. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Jiang, R.; Dong, B.; Fang, S.; Li, Q.; Lv, Z.; Chen, W. Phytohormone-Based Regulation of Trichome Development. Front. Plant Sci. 2021, 12, 734776. [Google Scholar] [CrossRef]
- Matías-Hernández, L.; Aguilar-Jaramillo, A.E.; Cigliano, R.A.; Sanseverino, W.; Pelaz, S. Flowering and Trichome Development Share Hormonal and Transcription Factor Regulation. J. Exp. Bot. 2016, 67, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, S.; Khan, A.R.; Gan, Y. TOE1/TOE2 Interacting with GIS to Control Trichome Development in Arabidopsis. Int. J. Mol. Sci. 2023, 24, 6698. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, D.; Khan, A.R.; Liu, B.; Wu, M.; Huang, L.; Wu, J.; Song, G.; Ni, H.; Ying, H.; et al. NbGIS Regulates Glandular Trichome Initiation through GA Signaling in Tobacco. Plant Mol. Biol. 2018, 98, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, D.; Hu, R.; Hua, C.; Ali, I.; Zhang, A.; Liu, B.; Wu, M.; Huang, L.; Gan, Y. AtGIS, a C2H2 Zinc-Finger Transcription Factor from Arabidopsis Regulates Glandular Trichome Development through GA Signaling in Tobacco. Biochem. Biophys. Res. Commun. 2017, 483, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Su, D.; Li, J.; Ying, S.; Deng, H.; He, X.; Zhu, Y.; Li, Y.; Chen, Y.; Pirrello, J.; et al. Overexpression of bHLH95, a Basic Helix–Loop–Helix Transcription Factor Family Member, Impacts Trichome Formation via Regulating Gibberellin Biosynthesis in Tomato. J. Exp. Bot. 2020, 71, 3450–3462. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, P.; Wang, B.; Yuan, F. The Roles of Trichome Development Genes in Stress Resistance. Plant Growth Regul. 2021, 95, 137–148. [Google Scholar] [CrossRef]
- Li, X.; Guo, C.; Ahmad, S.; Wang, Q.; Yu, J.; Liu, C.; Guo, Y. Systematic Analysis of MYB Family Genes in Potato and Their Multiple Roles in Development and Stress Responses. Biomolecules 2019, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, R.H.; Juenger, T.E.; Symonds, V.V. Natural Variation in GL1 and Its Effects on Trichome Density in Arabidopsis Thaliana. Mol. Ecol. 2012, 21, 3501–3515. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Pang, M.; Nah, G.; Shi, X.; Ye, W.; Stelly, D.M.; Chen, Z.J. miR828 and miR858 Regulate Homoeologous MYB2 Gene Functions in Arabidopsis Trichome and Cotton Fibre Development. Nat. Commun. 2014, 5, 3050. [Google Scholar] [CrossRef]
- Matías-Hernández, L.; Jiang, W.; Yang, K.; Tang, K.; Brodelius, P.E.; Pelaz, S. AaMYB1 and Its Orthologue AtMYB61 Affect Terpene Metabolism and Trichome Development in Artemisia annua and Arabidopsis thaliana. Plant J. 2017, 90, 520–534. [Google Scholar] [CrossRef]
- Zheng, K.; Tian, H.; Hu, Q.; Guo, H.; Yang, L.; Cai, L.; Wang, X.; Liu, B.; Wang, S. Ectopic Expression of R3 MYB Transcription Factor Gene OsTCL1 in Arabidopsis, but Not Rice, Affects Trichome and Root Hair Formation. Sci. Rep. 2016, 6, 19254. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-S.; Chen, M.; Li, L.-C.; Ma, Y.-Z. Functions and Application of the AP2/ERF Transcription Factor Family in Crop Improvement. J. Integr. Plant Biol. 2011, 53, 570–585. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Gao, D.; Xiong, Y.; Tang, X.; Xiao, X.; Wang, C.; Yu, S. Hairy Leaf 6, an AP2/ERF Transcription Factor, Interacts with OsWOX3B and Regulates Trichome Formation in Rice. Mol. Plant 2017, 10, 1417–1433. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, T.; Li, Y.; Liu, H.; Qin, W.; Wu, Z.; Peng, B.; Wang, X.; Yan, X.; Fu, X.; et al. AaWIN1, an AP2/ERF Protein, Positively Regulates Glandular Secretory Trichome Initiation in Artemisia annua. Plant Sci. 2023, 329, 111602. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Q.; Liu, M.; Bo, C.; Wang, X.; Ma, Q.; Cheng, B.; Cai, R. Overexpression of a Maize MYB48 Gene Confers Drought Tolerance in Transgenic Arabidopsis Plants. J. Plant Biol. 2017, 60, 612–621. [Google Scholar] [CrossRef]
- Yin, D.; Sun, D.; Han, Z.; Ni, D.; Norris, A.; Jiang, C.-Z. PhERF2, an Ethylene-Responsive Element Binding Factor, Plays an Essential Role in Waterlogging Tolerance of Petunia. Hortic. Res. 2019, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tian, Y.; Xu, J.; Fu, X.; Gao, J.; Wang, B.; Han, H.; Wang, L.; Peng, R.; Yao, Q. A Tomato ERF Transcription Factor, SlERF84, Confers Enhanced Tolerance to Drought and Salt Stress but Negatively Regulates Immunity against Pseudomonas syringae Pv. Tomato DC3000. Plant Physiol. Biochem. 2018, 132, 683–695. [Google Scholar] [CrossRef]
- Agarwal, P.; Mitra, M.; Banerjee, S.; Roy, S. MYB4 Transcription Factor, a Member of R2R3-Subfamily of MYB Domain Protein, Regulates Cadmium Tolerance via Enhanced Protection against Oxidative Damage and Increases Expression of PCS1 and MT1C in Arabidopsis. Plant Sci. 2020, 297, 110501. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D.G. Plant Pathogens and Integrated Defence Responses to Infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR Proteins in Pathogen Sensing and Host Defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef]
- Saintenac, C.; Zhang, W.; Salcedo, A.; Rouse, M.N.; Trick, H.N.; Akhunov, E.; Dubcovsky, J. Identification of Wheat Gene Sr35 That Confers Resistance to Ug99 Stem Rust Race Group. Science 2013, 341, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Periyannan, S.; Moore, J.; Ayliffe, M.; Bansal, U.; Wang, X.; Huang, L.; Deal, K.; Luo, M.; Kong, X.; Bariana, H.; et al. The Gene Sr33, an Ortholog of Barley Mla Genes, Encodes Resistance to Wheat Stem Rust Race Ug99. Science 2013, 341, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Cruz, C.M.; Paytuvi-Gallart, A.; Di Donato, A.; Sundesha, V.; Andolfo, G.; Aiese Cigliano, R.; Sanseverino, W.; Ercolano, M.R. PRGdb 3.0: A Comprehensive Platform for Prediction and Analysis of Plant Disease Resistance Genes. Nucleic Acids Res. 2018, 46, D1197–D1201. [Google Scholar] [CrossRef]
- Liu, P.-L.; Du, L.; Huang, Y.; Gao, S.-M.; Yu, M. Origin and Diversification of Leucine-Rich Repeat Receptor-like Protein Kinase (LRR-RLK) Genes in Plants. BMC Evol. Biol. 2017, 17, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, T.; Wang, W.; Hu, H.; Wei, Q.; Bao, C. Investigation of Evolutionary and Expressional Relationships in the Function of the Leucine-Rich Repeat Receptor-like Protein Kinase Gene Family (LRR-RLK) in the Radish (Raphanus sativus L.). Sci. Rep. 2019, 9, 6937. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, P.A.; Shan, L.; He, P. Plant Cell Surface Molecular Cypher: Receptor-like Proteins and Their Roles in Immunity and Development. Plant Sci. 2018, 274, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ding, Y.; Jiang, Q.; Wang, F.; Sun, J.; Zhu, C. The Role of Receptor-like Protein Kinases (RLKs) in Abiotic Stress Response in Plants. Plant Cell Rep. 2017, 36, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Sensing the Environment: Key Roles of Membrane-Localized Kinases in Plant Perception and Response to Abiotic Stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Jose, J.; Ghantasala, S.; Roy Choudhury, S. Arabidopsis Transmembrane Receptor-Like Kinases (RLKs): A Bridge between Extracellular Signal and Intracellular Regulatory Machinery. Int. J. Mol. Sci. 2020, 21, 4000. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroid Signal Transduction: From Receptor Kinase Activation to Transcriptional Networks Regulating Plant Development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef]
- Tong, H.; Chu, C. Functional Specificities of Brassinosteroid and Potential Utilization for Crop Improvement. Trends Plant Sci. 2018, 23, 1016–1028. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional Regulators of Plant Growth, Development, and Stress Responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Imkampe, J.; Halter, T.; Huang, S.; Schulze, S.; Mazzotta, S.; Schmidt, N.; Manstretta, R.; Postel, S.; Wierzba, M.; Yang, Y.; et al. The Arabidopsis Leucine-Rich Repeat Receptor Kinase BIR3 Negatively Regulates BAK1 Receptor Complex Formation and Stabilizes BAK1. Plant Cell 2017, 29, 2285–2303. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Ni, Y.; Jia, M.; Liu, H. Comprehensive Analysis of Sesame LRR-RLKs: Structure, Evolution and Dynamic Expression Profiles under Macrophomina Phaseolina Stress. Front. Plant Sci. 2024, 15, 1334189. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yin, W.; Liu, D.; Tong, H. Regulation of Brassinosteroid Signaling and Salt Resistance by SERK2 and Potential Utilization for Crop Improvement in Rice. Front. Plant Sci. 2020, 11, 621859. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zuo, S.; Schwessinger, B.; Chern, M.; Canlas, P.E.; Ruan, D.; Zhou, X.; Wang, J.; Daudi, A.; Petzold, C.J.; et al. An XA21-Associated Kinase (OsSERK2) Regulates Immunity Mediated by the XA21 and XA3 Immune Receptors. Mol. Plant 2014, 7, 874–892. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Dai, C.; Lee, M.M.; Kwak, J.M.; Nam, K.H. BRI1-Associated Receptor Kinase 1 Regulates Guard Cell ABA Signaling Mediated by Open Stomata 1 in Arabidopsis. Mol. Plant 2016, 9, 447–460. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Mao, L.; Zhou, Y.; Sun, Y.; Lv, J.; Deng, M.; Liu, Z.; Yang, B. Transcriptome Analysis Reveals Key Genes Involved in Trichome Formation in Pepper (Capsicum annuum L.). Plants 2024, 13, 1090. https://doi.org/10.3390/plants13081090
Shen Y, Mao L, Zhou Y, Sun Y, Lv J, Deng M, Liu Z, Yang B. Transcriptome Analysis Reveals Key Genes Involved in Trichome Formation in Pepper (Capsicum annuum L.). Plants. 2024; 13(8):1090. https://doi.org/10.3390/plants13081090
Chicago/Turabian StyleShen, Yiyu, Lianzhen Mao, Yao Zhou, Ying Sun, Junheng Lv, Minghua Deng, Zhoubin Liu, and Bozhi Yang. 2024. "Transcriptome Analysis Reveals Key Genes Involved in Trichome Formation in Pepper (Capsicum annuum L.)" Plants 13, no. 8: 1090. https://doi.org/10.3390/plants13081090