
Neuroprotective Effects of Chemical Constituents of Leaves of Euonymus hamiltonianus Wall.


 , ,
, ,
Abstract

1. Introduction
2. Results and Discussion
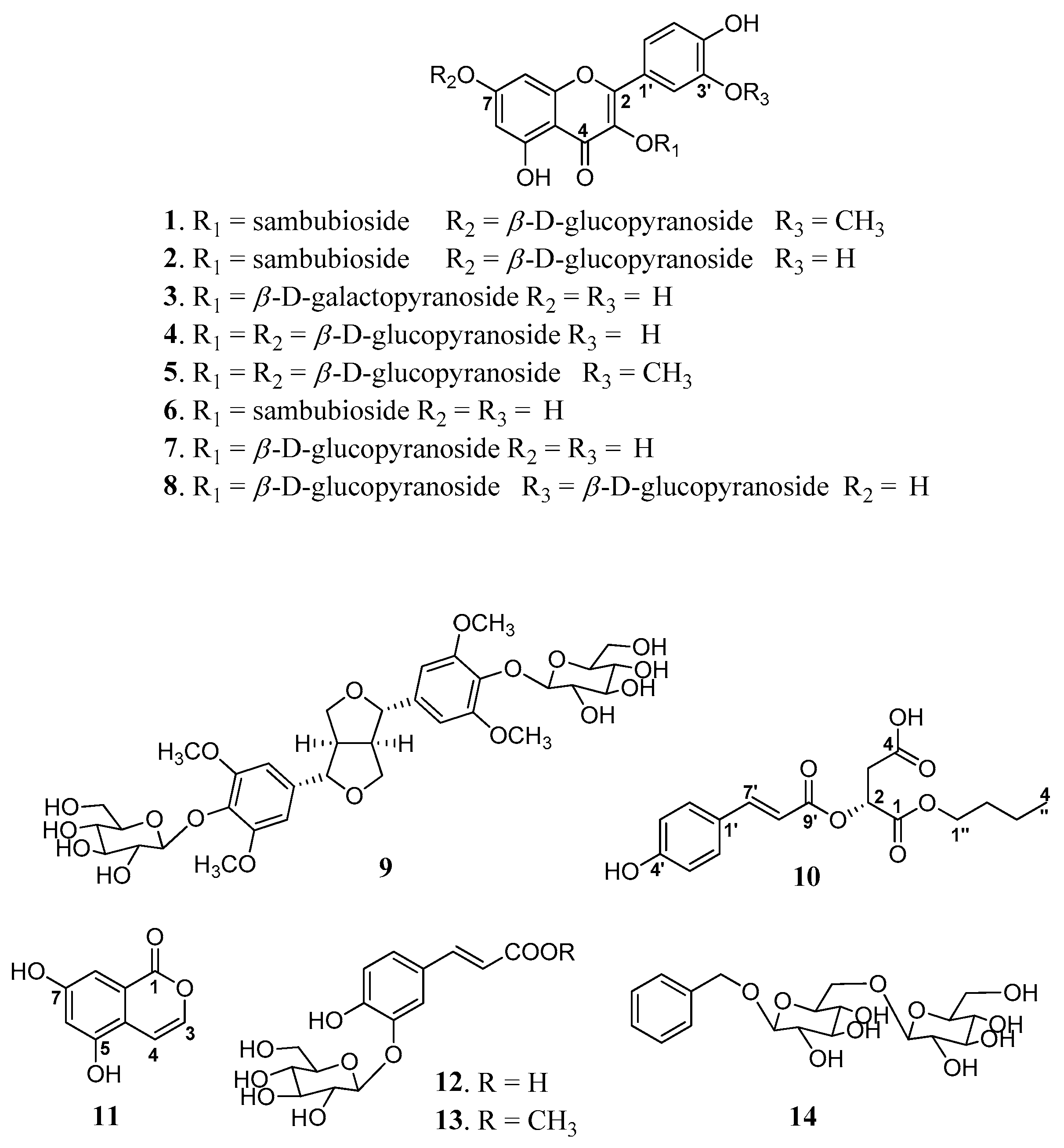
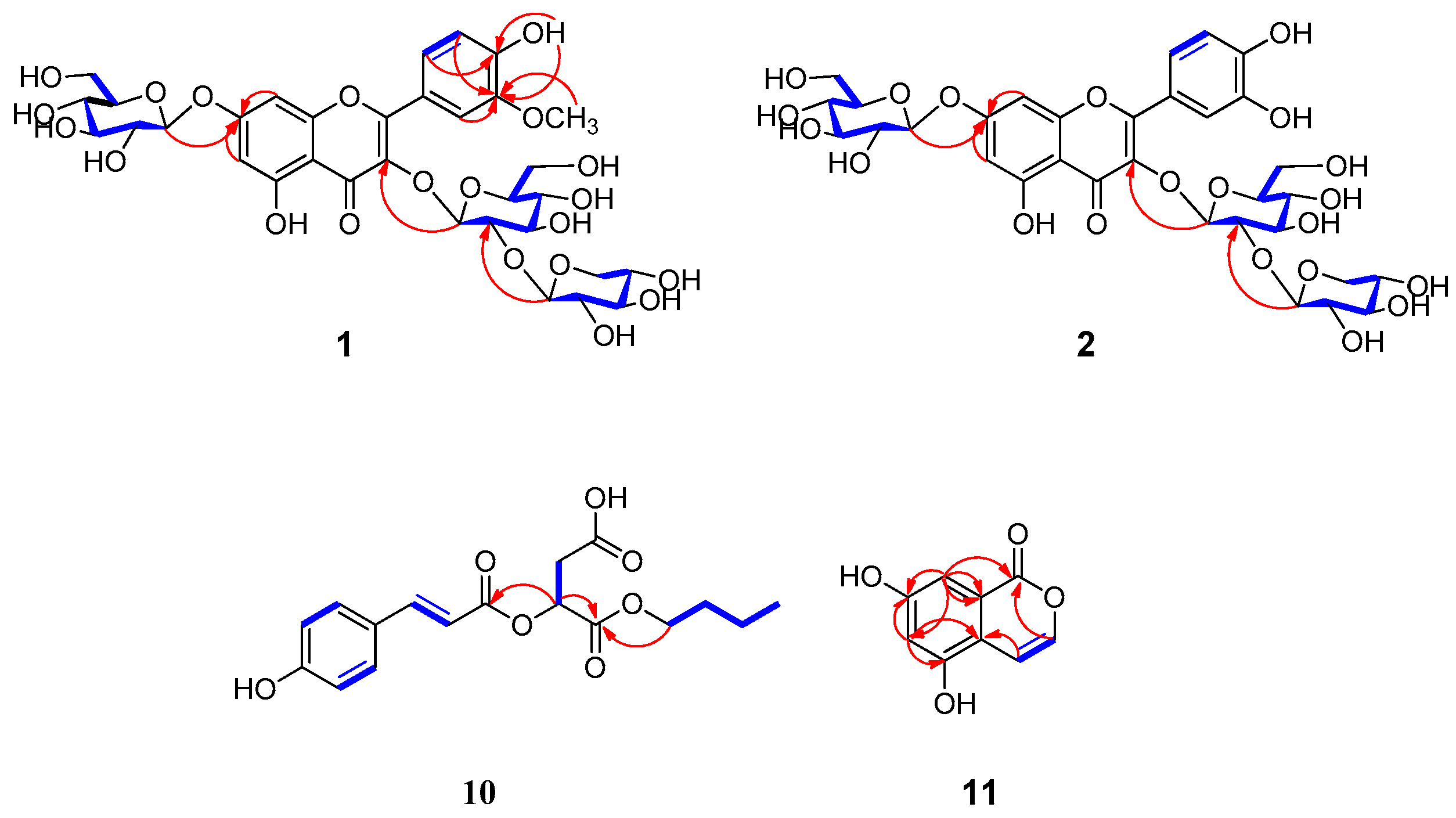
2.1. Structural Determination of Compounds 1, 2, 10, and 11
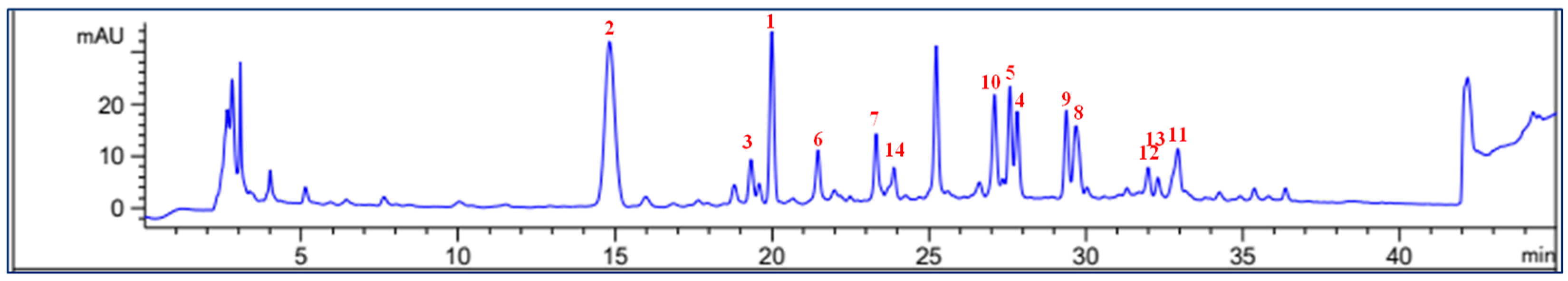
2.2. Chemical Profiling of Phytochemicals 1–14
2.3. Inhibition of the Aβ Aggregation of Compound 13
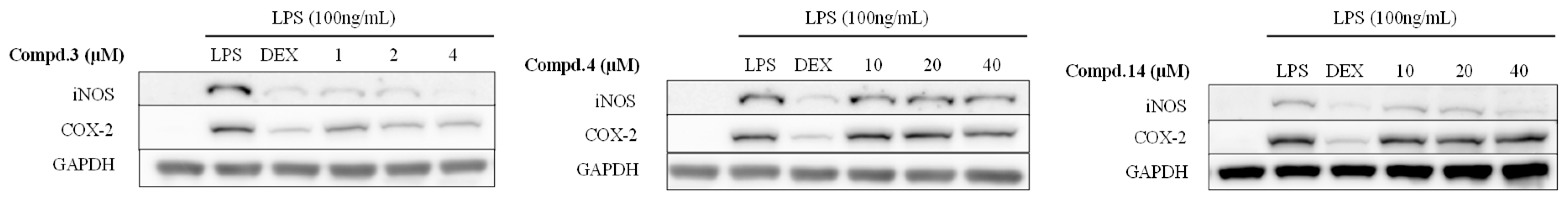
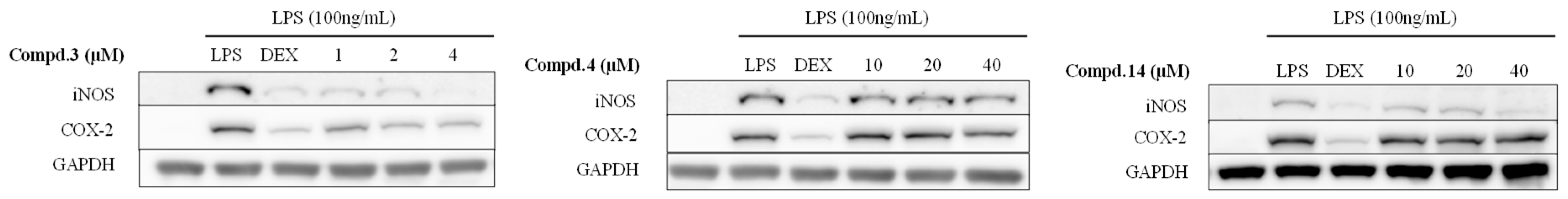
2.4. Anti-Inflammation Effect of the Isolated Compounds 3, 4, and 14
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Materials
3.3. Extraction and Isolation
3.4. Euonymusin A (1)
3.5. Euonymusin D (2)
3.6. Euonymusin B (10)
3.7. Euonymusin C (11)
3.8. Hydrolysis of 1
3.9. Aβ Assay
3.10. Cell Cultures and MTT Assay for Cell Viability
3.11. Determination of NO Production and the Cell Viability Assay
3.12. Western Blot Analysis
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nix, N.M.; Price, A. Acute Myeloid Leukemia: An Ever-Changing Disease. J. Adv. Pract. Oncol. 2019, 10, 4–8. [Google Scholar]
- Ransohoff, R.M. How neuroinflammation contributes to neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Murfphy, M.P.; LeVine, H. Alzheimer’s disease and the amyloid-beta peptide. J. Alzheimers Dis. 2010, 19, 311–323. [Google Scholar] [CrossRef]
- Cole, S.L.; Vassar, R. The Role of Amyloid Precursor Protein Processing by BACE1, the β-Secretase, in Alzheimer Disease Pathophysiology. J. Biol. Chem. 2008, 293, 29621–29625. [Google Scholar] [CrossRef] [PubMed]
- Gandy, S. The role of cerebral amyloid beta accumulation in common forms of Alzheimer’s disease. J. Clin. Investig. 2005, 115, 1121–1129. [Google Scholar]
- Silva, T.; Reis, J.; Teixeira, J.; Borges, F. Alzheimer’s disease, enzyme targets and drug discovery struggles: From natural products to drug prototypes. Ageing Res. Rev. 2014, 15, 116–145. [Google Scholar] [CrossRef]
- Jung, W.K.; Heo, S.J.; Jeon, Y.J.; Lee, C.M.; Park, Y.M.; Byun, H.G.; Choi, Y.H.; Park, S.G.; Choi, I.W. Inhibitory effects and molecular mechanism of dieckol isolated from marine brown alga on COX-2 and iNOS in microglial cells. J. Agri. Food Chem. 2009, 57, 4439–4446. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef]
- Sahebnasagh, A.; Eghbali, S.; Saghafi, F.; Sureda, A.; Avan, R. Neurohormetic phytochemicals in the pathogenesis of neurodegenerative diseases. Immun. Ageing 2022, 19, 81–90. [Google Scholar] [CrossRef]
- Kumar, G.P.; Khanum, F. Neuroprotective potential of phytochemicals. Pharmacogn. Rev. 2012, 6, 81–90. [Google Scholar] [CrossRef]
- Poudel, P.; Thapa, R.; Devkota, H.P.; Yahara, S.; Jamarkattel-Pandit, N. Chemical analysis, antidiabetic and antioxidant activity of Euonymus hamiltonianus Wall. Indian J. Pharmacol. 2013, 45, S171–S172. [Google Scholar]
- Sharma, A.; Sati, S.C.; Sati, O.P.; Sati, M.D.; Kothiyal, S.K. Genus Euonymus: Chemical and Pharmacological Perception. Mini-Rev. Org. Chem. 2012, 9, 341–351. [Google Scholar] [CrossRef]
- Ishtiwata, H.; Shizuri, Y.; Yamada, K. Three sesquiterpene alkaloids from Euonymus alatus forma Striatus. Phytochemistry 1983, 22, 2839–2841. [Google Scholar] [CrossRef]
- Hohmann, J. New sesquiterpene polyesters from Euonymus species. J. Nat. Prod. 1995, 58, 1192–1199. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Mehrotra, B.N. Compendium of Indian Medicinal Plants; Central Drug Research Institute and Publications & Information Directorate: New Delhi, India, 1994; Volume 5, pp. 388–389. [Google Scholar]
- Tantray, M.A.; Shawl, A.S.; Khuroo, M.A.; Bhat, B.A. Two new coumarins from Euonymus hamiltonianus. Fitoterapia 2008, 79, 234–235. [Google Scholar] [CrossRef]
- Tantray, M.A.; Shawl, A.S.; Arora, B.S.; Purinima, N.; Ahmad, K.; Khuroo, M.A. Glutinane triterpenes from the stem bark of Euonymus hamiltonianus. Chem. Nat. Compd. 2009, 45, 377–380. [Google Scholar] [CrossRef]
- Tantry, M.A.; Khuroo, M.A.; Shawl, A.S.; Najar, M.H.; Khan, I.A. Dihydro-β-agarofuran sesquiterpene pyridine alkaloids from the seeds of Euonymus hamiltonianus. J. Saudi Chem. Soc. 2016, 20, S323–S327. [Google Scholar] [CrossRef]
- Pant, R.R. Chemical constituents of Euonymus hamitonianus Wall. J. Nepal Chem. Soc. 1993, 12, 339–343. [Google Scholar]
- Sasaki, K.; Takahashi, T. A flavonoid from Brassica rapa flower as the UV-absorbing nectar guide. Phytochemistry 2002, 61, 339–343. [Google Scholar] [CrossRef]
- Tran, H.N.K.; Youn, U.J.; Kim, J.A.; Chae, H.; Kim, S.; Min, B.S. Glycerols and fatty acids isolated from Micractinium sp. KSF0031. Biochem. Syst. Ecol. 2020, 89, 104000. [Google Scholar]
- Gluchofffiasson, K.; Fiasson, J.L.; Favrebonvin, J. Quercetin glycosides from antarctic Ranunculus species. Phytochemistry 1994, 37, 1629–1633. [Google Scholar] [CrossRef]
- Lee, K.H.; Cho, J.Y.; Lee, H.J.; Ma, Y.K.; Kwon, J.; Park, S.H.; Lee, S.H.; Cho, J.A.; Kim, W.S.; Park, K.H.; et al. Hydroxycinnamoylmalic Acids and Their Methyl Esters from Pear (Pyrus pyrifolia Nakai) Fruit Peel. J. Agric. Food Chem. 2011, 59, 10124–10128. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Jeon, H.Y.; Lee, K.Y.; Kim, S.H.; Ma, C.J.; Sung, S.H.; Lee, H.S.; Park, M.J.; Kim, Y.C. Inhibitory constituents of Euscaphis japonica on lipopolysaccharide-induced nitric oxide production in BV2 microglia. Planta Med. 2007, 73, 782–786. [Google Scholar] [CrossRef]
- Sakamoto, F.; Ikeda, S.; Tsukamoto, G. Studies on prodrugs. I. preparation and characterization of acyloxyallylester of ampicillin. Chem. Pharm. Bull. 1983, 31, 2698–2707. [Google Scholar] [CrossRef]
- Choi, S.J.; Tai, B.H.; Cuong, N.M.; Kim, Y.H.; Jang, H.D. Antioxidative and anti-inflammatory effect of quercetin and its glycosides isolated from mampat (Cratoxylum formosum). Food Sci. Biotechnol. 2012, 21, 587–595. [Google Scholar] [CrossRef]
- Yokozawa, T.; Kim, H.Y.; Cho, E.J.; Choi, J.S.; Chung, H.Y. Antioxidant effects of isorhamnetin 3,7-di-O-β-D-glucopyranoside isolated from mustard leaf (Brassica juncea) in rats with streptozotocin-induced diabetes. J. Agric. Food Chem. 2002, 50, 5490–5495. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.P.; Liu, S.Y.; Fei, D.Q.; Zhang, M.; Zhang, Y.X. Effect of quercetin-3-O-sambubioside isolated from Eucommia ulmoides male flowers on spontaneous activity and convulsion rate in mice. Planta Med. 2014, 80, 974–977. [Google Scholar] [CrossRef] [PubMed]
- Su, X.D.; Jang, H.J.; Li, H.X.; Kim, Y.H.; Yang, S.Y. Identification of potential inflammatory inhibitors from Aster tataricus. Bioorg. Chem. 2019, 92, 103208. [Google Scholar] [CrossRef]
- Ceska, O.; Styles, E.D. Flavonoids from Zea mays pollen. Phytochemistry 1984, 23, 1822–1823. [Google Scholar] [CrossRef]
- Kobayashi, H.; Karasaw, H.; Miyase, T. Studies on the constituents of Cistanchis herba. V. Isolation and structures of two new phenylpropanoid glycosides, cistanosides E and F. Chem. Pharm. Bull. 1985, 33, 1452–1457. [Google Scholar] [CrossRef]
- Limjiasahapong, S.; Tuchinda, P.; Reutrakul, V.; Pohmakotr, M.; Akkarawongsapat, R.; Limthongkul, J.; Napaswad, C.; Nuntasaen, N. Anti-HIV-1 activities and chemical constituents from leaves and twigs of Santisukia pagetii (Bignoniaceae). Nat. Prod. Commun. 2018, 13, 1449–1452. [Google Scholar]
- DeRosa, S.; DeGiulio, A.; Tommonaro, G. Aliphatic and aromatic glycosides from the cell cultures of Lycopersicon esculentum. Phytochemistry 1996, 42, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Magdalena, B. Influence of extraction methods on stability of flavonoids. J. Chromatogr. A 2011, 1218, 2505–2512. [Google Scholar]
- Liu, X.M.; Liu, Y.; Shan, C.H.; Yang, X.Q.; Zhang, Q.; Xu, N.; Xu, L.Y.; Song, W. Effects of five extraction methods on total content, composition, and stability of flavonoids in jujube. Food Chem. X 2022, 14, 100287. [Google Scholar] [CrossRef] [PubMed]
- Pagano, K.; Tomaselli, S.; Molinari, H.; Ragona, L. Natural compounds as inhibitors of Aβ peptide aggregation: Chemical requirements and molecular mechanisms. Front. Neurosci. 2020, 14, 619667. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective effects of quercetin in Alzheimer’s disease. Biomolecules 2020, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Askarova, S.; Yang, X.; Lee, J.C.M. Impacts of membrane biophysics in Alzheimer’s disease: From amyloid precursor protein processing to Aβ peptide-induced membrane changes. Int. J. Alzheimers. Dis. 2011, 2011, 134971. [Google Scholar] [CrossRef]
- Li, X.; Cui, J.; Yu, Y.; Li, W.; Hou, Y.; Wang, X.; Qin, D.; Zhao, C.; Yao, X.; Zhao, J.; et al. Traditional Chinese Nootropic Medicine Radix Polygalae and Its Active Constituent Onjisaponin B Reduce β-Amyloid Production and Improve Cognitive Impairments. PLoS ONE 2016, 11, e0151147. [Google Scholar] [CrossRef]
- Rehman, S.U.; Ali, T.; Alam, S.I.; Ullah, R.; Zeb, A.; Lee, K.W.; Rutten, B.P.; Kim, M.O. Ferulic acid rescues LPS-induced neurotoxicity via modulation of the TLR4 receptor in the mouse hippocampus. Mol. Neurobiol. 2019, 56, 2774–2790. [Google Scholar] [CrossRef]
- Chu, F.; Shi, M.; Zheng, C.; Shen, D.; Zhu, J.; Zheng, X.; Cui, L. The roles of macrophages and microglia in multiple sclerosis and experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2018, 318, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.; Koliada, A.; Lushchak, O. Neuroinflammation in pathogenesis of Alzheimer’s disease: Phytochem-icals as potential therapeutics. Mech. Ageing Dev. 2020, 189, 111259. [Google Scholar] [CrossRef] [PubMed]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the War Against Inflammation With Natural Products. Front Pharmacol. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.E. Recent development on COX-2 inhibitors as promising an-ti-inflammatory agents: The past 10 years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef]
- Lee, S.B.; Yang, S.Y.; Thao, N.P.; Seo, D.G.; Kim, S.; Ma, C.T.; Park, S.Y.; Kim, Y.H.; Yang, H.O. Protective effects of compounds from Cimicifuga dahurica against amyloid beta production in vitro and scopolamine-induced memory impairment in vivo. J. Nat. Prod. 2020, 83, 223–230. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 a | 2 a | 10 a | 11 b | ||||
|---|---|---|---|---|---|---|---|---|
| δH (J in Hz) | δC | δH (J in Hz) | δC | δH (J in Hz) | δC | δH (J in Hz) | δC | |
| 1 | 169.0 | 183.4 | ||||||
| 2 | 156.4 | 156.1 | 5.37, dd (8.0, 4.0) | 67.9 | ||||
| 3a | 133.2 | 133.3 | 2.86, dd (12.0, 4.5) | 35.7 | 7.97, d (5.6) | 158.2 | ||
| 3b | 2.79, dd (12.0, 3.5) | |||||||
| 4 | 177.6 | 177.6 | 170.7 | 6.19, ovl c | 111.6 | |||
| 4a | 105.7 | 105.6 | 159.9 | |||||
| 5 | 160.9 | 161.0 | 163.5 | |||||
| 6 | 6.44, d (2.0) | 99.3 | 6.42, d (2.0) | 99.3 | 6.19, ovl c | 100.2 | ||
| 7 | 162.9 | 162.8 | 166.2 | |||||
| 8 | 6.82, d (2.0) | 94.6 | 6.76, d (2.0) | 94.3 | 6.33, d (1.6) | 95.1 | ||
| 8a | 156.0 | 156.0 | 106.6 | |||||
| 1′ | 121.0 | 121.1 | 124.9 | |||||
| 2′ | 7.86, d (2.0) | 113.2 | 7.59, d (2.5) | 115.3 | 7.59, d (8.5) | 130.5 | ||
| 3′ | 147.1 | 145.0 | 6.79, d (8.5) | 115.5 | ||||
| 4′ | 149.8 | 148.8 | 160.3 | |||||
| 5′ | 6.93, d (8.5) | 115.3 | 6.86, d (8.0) | 116.4 | 6.79, d (8.5) | 115.5 | ||
| 6′ | 7.65, dd (8.5, 2.0) | 122.8 | 7.67, dd (8.0, 2.5) | 122.1 | 7.59, d (8.5) | 130.5 | ||
| 7′ | 7.60, d (16.0) | 145.9 | ||||||
| 8′ | 6.46, d (16.0) | 112.8 | ||||||
| 9′ | 165.8 | |||||||
| 1″ | 5.76, d (7.0) | 98.1 | 5.73, d (7.0) | 97.9 | 4.10, m | 64.4 | ||
| 2″ | 3.48, d (7.0) | 81.6 | 3.49, d (7.0) | 82.0 | 1.54, m | 29.7 | ||
| 3″ | 3.13, m | 77.6 | 3.10, m | 77.8 | 1.30, m | 18.2 | ||
| 4″ | 3.05, dd (7.0, 3.0) | 73.9 | 3.29, d (8.0) | 74.1 | 0.86, t (7.0) | 13.3 | ||
| 5″ | 3.25, m | 73.2 | 3.14, m | 73.2 | ||||
| 6″a | 3.55, m | 60.5 | 3.70, m | 60.7 | ||||
| 6″b | 3.35, m | 3.53, d (11.5) | ||||||
| 1‴ | 4.62, d (7.5) | 104.2 | 4.58, d (7.5) | 104.7 | ||||
| 2‴ | 3.44, d (8.0) | 77.3 | 3.06, d (7.5) | 77.2 | ||||
| 3‴ | 3.00, d (8.0) | 76.2 | 3.14, m | 76.3 | ||||
| 4‴ | 3.23, m | 69.4 | 3.24, d (9.0) | 69.4 | ||||
| 5‴a | 3.66, dd (11.0, 5.0) | 65.6 | 3.67, m | 65.8 | ||||
| 5‴b | 2.98, d (11.0) | 3.03, m | ||||||
| 1⁗ | 5.08, d (7.5) | 99.8 | 5.10, d (7.5) | 99.7 | ||||
| 2⁗ | 3.12, m | 69.6 | 3.27, d (7.5) | 69.6 | ||||
| 3⁗ | 3.28, m | 76.5 | 3.10, m | 76.5 | ||||
| 4⁗ | 3.11, m | 77.1 | 3.46, d (8.0) | 76.9 | ||||
| 5⁗ | 3.16, m | 69.5 | 3.11, m | 69.5 | ||||
| 6⁗a | 3.69, m | 60.7 | 3.44, m | 60.7 | ||||
| 6⁗b | 3.46, d (7.0) | 3.31, m | ||||||
| 3′-OCH3 | 3.85, s | 55.8 | ||||||
| 5-OH | 12.60, s | 12.69, brs | ||||||
| 4′-OH | 9.95, brs | |||||||
| 3″-OH | 4.46, d (5.5) | |||||||
| 4″-OH | 5.20, brs | |||||||
| 6″-OH | 4.45, d (5.5) | |||||||
| 2‴-OH | 4.66, d (6.0) | |||||||
| 3‴-OH | 5.54, d (4.0) | |||||||
| 4‴-OH | 5.13, brs | |||||||
| 2⁗-OH | 5.02, m | |||||||
| 3⁗-OH | 5.45, d (4.5) | |||||||
| 4⁗-OH | 5.49, d (4.0) | |||||||
| 6⁗-OH | 4.65, d (5.5) | |||||||
| OH | 4.93, brs | |||||||
| Compounds | IC50 Value (µM) a | ||
|---|---|---|---|
| Aβ42 | NO | Cytotoxicity | |
| 1 | - | >100 | >100 |
| 2 | >100 | >100 | >100 |
| 3 | - | 16.33 ± 2.94 | >100 |
| 4 | - | 14.38 ± 8.68 | >100 |
| 5 | - | >100 | >100 |
| 6 | >100 | >100 | >100 |
| 7 | - | >100 | >100 |
| 8 | - | >100 | >100 |
| 9 | - | >100 | >100 |
| 10 | - | >100 | >100 |
| 11 | - | >100 | >100 |
| 12 | >100 | >100 | >100 |
| 13 | 65.43 ± 6.71 | >100 | >100 |
| 14 | - | 17.44 ± 2.77 | >100 |
| Justicidine A b | 1.0 ± 0.043 | ||
| Dexamethasone b | 1.24 ± 0.104 | >100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, H.N.K.; Yu, J.S.; Huang, T.; Lee, G.; Choi, H.S.; Yang, H.O. Neuroprotective Effects of Chemical Constituents of Leaves of Euonymus hamiltonianus Wall. Plants 2024, 13, 1094. https://doi.org/10.3390/plants13081094
Tran HNK, Yu JS, Huang T, Lee G, Choi HS, Yang HO. Neuroprotective Effects of Chemical Constituents of Leaves of Euonymus hamiltonianus Wall. Plants. 2024; 13(8):1094. https://doi.org/10.3390/plants13081094
Chicago/Turabian StyleTran, Huynh Nguyen Khanh, Jae Sik Yu, Tianqi Huang, Gakyung Lee, Hyo Sun Choi, and Hyun Ok Yang. 2024. "Neuroprotective Effects of Chemical Constituents of Leaves of Euonymus hamiltonianus Wall." Plants 13, no. 8: 1094. https://doi.org/10.3390/plants13081094
APA StyleTran, H. N. K., Yu, J. S., Huang, T., Lee, G., Choi, H. S., & Yang, H. O. (2024). Neuroprotective Effects of Chemical Constituents of Leaves of Euonymus hamiltonianus Wall. Plants, 13(8), 1094. https://doi.org/10.3390/plants13081094