Regeneration of Genetically Stable Plants from in Vitro Vitrified Leaves of Different Carnation Cultivars


Abstract
:1. Introduction
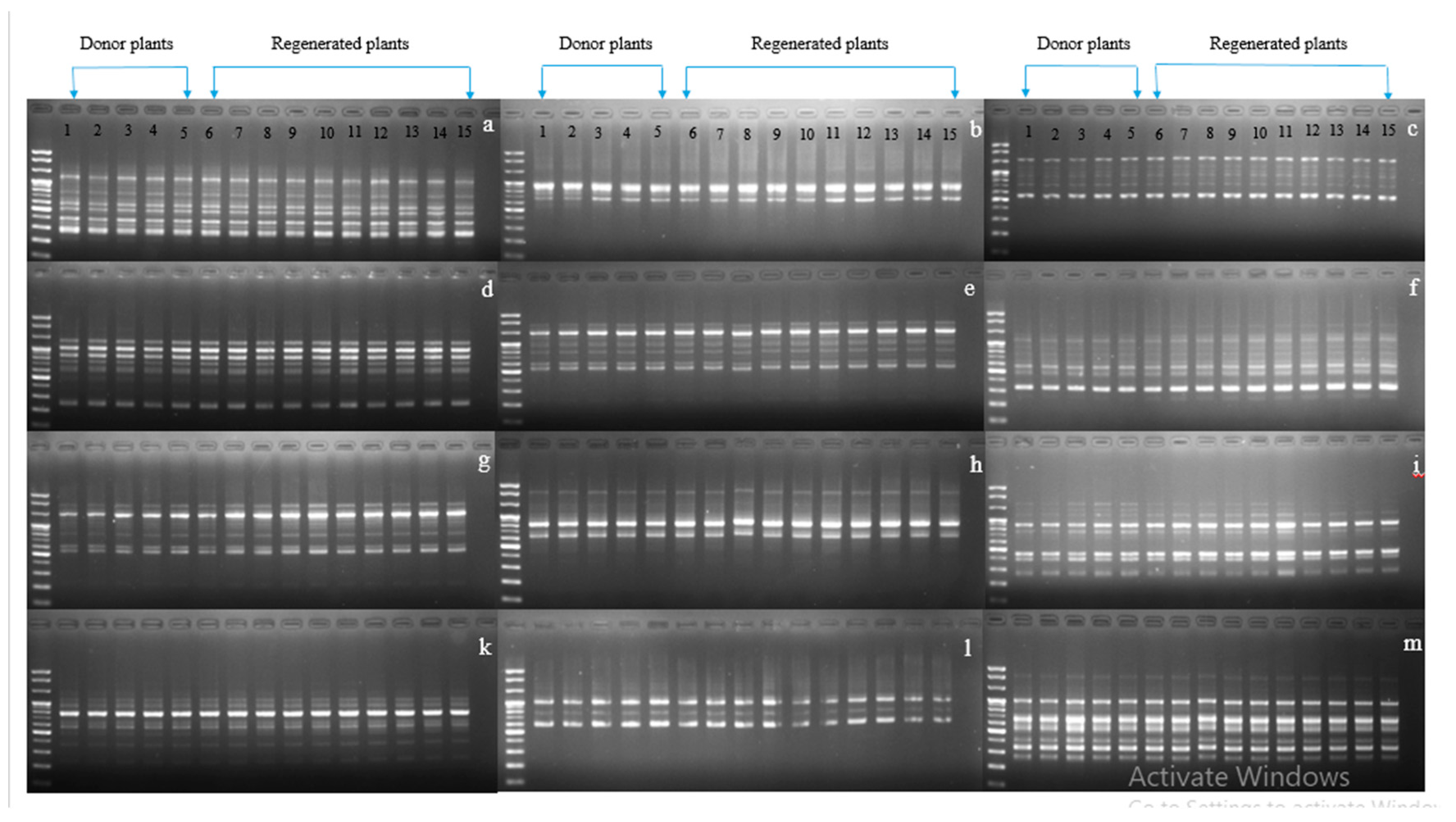
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Plant Materials
4.2. Induction of Normal and Vitrified Leaves Development
4.3. Shoot Induction from Normal and Vitrified Leaves
4.4. Plant Growth and Rooting
4.5. Acclimatization of in vitro Regenerated Plants
4.6. Detection of Genetic Variation by RAPD
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kumar, S.; Kumari, R.; Baheti, T.; Thakur, M.; Ghani, M. Plant regeneration from axillary bud, callus and somatic embryo in carnation (Dianthus caryophyllus) and assessment of genetic fidelity using RAPD-PCR analysis. Indian J. Agric. Sci. 2016, 86, 1482–1488. [Google Scholar]
- Shiba, T.; Mii, M. Agrobacterium tumefaciens- mediated transformation of highly regenerable cell suspension cultures in Dianthus acicularis. J. Hortic. Sci. Biotechnol. 2005, 80, 393–398. [Google Scholar] [CrossRef]
- Van Altvorst, A.C.; Rikesen, T.; Koehorst, H.; Dons, J.J.M. Transgenic carnations obtained by Agrobacterium tumefaciens mediated of leaf explants. Transgenic. Res. 1995, 4, 105–113. [Google Scholar] [CrossRef]
- Lu, C.-Y.; Nugent, G.; Wardley-Richardson, T.; Chandler, S.; Young, R.; Dalling, M.J. Agrobacterium-Mediated Transformation of Carnation (Dianthus caryophyllus L.). Nat. Biotechnol. 1991, 9, 864–868. [Google Scholar] [CrossRef]
- Meng, L.-S.; Song, J.-P.; Sun, S.-B.; Wang, C.-Y. The ectopic expression of PttKN1 gene causes pleiotropic alternation of morphology in transgenic carnation (Dianthus caryophyllus L.). Acta Physiol. Plant. 2009, 31, 1155–1164. [Google Scholar] [CrossRef]
- Ovadis, M.; Zuker, A.; Tzfira, T.; Ahroni, A.; Shklarman, E.; Scovel, G.; Itzhaki, H.; Ben-Meir, H.; Vainstein, A. Generation of transgenic carnation plants with novel characteristics by combining microprojectile bombardment with Agrobacterium tumefaciens transformation. In Plant biotechnology and in vitro biology in the 21 st; Springer: Dordrecht, The Netherlands, 1999; Volume 36, pp. 189–192. [Google Scholar]
- Zuker, A.; Tzfira, T.; Scovel, G.; Ovadis, M.; Shklarman, E.; Itzhaki, H.; Vainstein, A. RolC-Transgenic Carnation with Improved Horticultural Traits: Quantitative and Qualitative Analyses of Greenhouse-grown Plants. J. Am. Soc. Hortic. Sci. 2001, 126, 13–18. [Google Scholar] [CrossRef]
- Casanova, E.; Moysset, L.; Trillasmava, M.I. Effects ofagar concentration and vessel closure on the organogenesis andhyperhydricity of adventitious carnation shoots. BiologiaPlantarum 2008, 52, 1–8. [Google Scholar]
- Gutierrez-Miceli, F.A.; Arias, L.; Juárez-Rodríguez, N.; Abud-Archila, M.; Amaro-Reyes, A.; Dendooven, L. Optimization of growth regulators and silver nitrate for micropropagation of Dianthus caryophyllus L. with the aid of a response surface experimental design. Vitr. Cell. Dev. Boil. Anim. 2009, 46, 57–63. [Google Scholar] [CrossRef]
- Yantcheva, A.; Vlahova, M.; Antanassov, A. Direct somatic embryogenesis and plant regeneration of carnation (Dianthos caryophyllus L.). Plant Cell Rep. 1998, 18, 148–153. [Google Scholar] [CrossRef]
- Karami, O.; Esna-Ashari, M.; Piri, K.; Almasi, P. Efficientregeneration of carnation (Dianthus caryophyllus L.) via somatic embryogenesis. Propag. Ornam. Plants 2007, 7, 3–8. [Google Scholar]
- Mante, S.; Evans, D.A.; Sharp, W.R.; Ammirato, P.V.; Yamada, Y. Handbook of Plant Cell Culture. Brittonia 1985, 37, 21. [Google Scholar] [CrossRef]
- Ziv, M. Vitrification: Morphological and physiological disorder of in vitro plants. In Micropropagation: Technology and Application; Debergh, P.C., Zimmerman, R.H., Eds.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 1991; pp. 45–69. [Google Scholar]
- Kevers, C.; Gaspar, T. Vitrification of carnation invitro: Changes in water content, extracellular space, air volume, and ion levels. Physiol. Vegétale Paris 1986, 24, 647–653. [Google Scholar]
- Phan, C.; Letouze, R. A comparative study of chlorophyll, phenolic and protein contents, and of hydroxycinnamate:CoA ligase activity of normal and ‘vitreous’ plants (Prunus avium L.) obtained in vitro. Plant Sci. Lett. 1983, 31, 323–327. [Google Scholar] [CrossRef]
- Böttcher, I.; Zoglauer, K.; Göring, H. Induction and reversion of vitrification of plants cultured in vitro. Physiol. Plant. (København 1948) 1988, 72, 560–564. [Google Scholar]
- Firoozabady, E.; Moy, Y.; Tucker, W.; Robinson, K.; Gutterson, N. Efficient transformation and regeneration of carnation cultivars usingAgrobacterium. Mol. Breed. 1995, 1, 283–293. [Google Scholar] [CrossRef]
- Kantia, A.; Kothari, S. High efficiency adventitious shoot bud formation and plant regeneration from leaf explants of Dianthus chinensis L. Sci. Hortic. 2002, 96, 205–212. [Google Scholar] [CrossRef]
- Arif, M.; Rauf, S.; Din, A.U.; Rauf, M.; Afrasiab, H. High Frequency Plant Regeneration from Leaf Derived Callus of Dianthus caryophyllus L. Am. J. Plant Sci. 2014, 5, 2454–2463. [Google Scholar] [CrossRef] [Green Version]
- Thakur, K.; Kanwar, K. In Vitro Plant Regeneration by Organogenesis from Leaf Callus of Carnation, Dianthus caryophyllus L. cv.‘Master’. Proceedings of the National Academy of Sciences. India Sect. B Biol. Sci. 2018, 88, 1147–1155. [Google Scholar]
- Muneer, S.; Park, G.Y.; Jeong, B.R. Red and Blue Light Emitting Diodes (LEDs) Participate in Mitigation of Hyperhydricity in In Vitro-Grown Carnation Genotypes (Dianthus Caryophyllus). J. Plant Growth Regul. 2018, 37, 370–379. [Google Scholar] [CrossRef]
- Sivanesan, I.; Song, J.Y.; Hwang, S.J.; Jeong, B.R. Micropropagation of Cotoneaster wilsonii Nakai—a rare endemic ornamental plant. Plant Cell, Tissue Organ Cult. (PCTOC) 2010, 105, 55–63. [Google Scholar] [CrossRef]
- Salisbury, F.; Ross, C. Plant physiology fourth edition. Respiration; Wadsworth Publishing Company: Belmont, CA, USA, 1992; pp. 266–288. [Google Scholar]
- Ahmadian, M.; Babaei, A.; Shokri, S.; Hessami, S. Micropropagation of carnation ( Dianthus caryophyllus L.) in liquid medium by temporary immersion bioreactor in comparison with solid culture. J. Genet. Eng. Biotechnol. 2017, 15, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Kevers, C.; Franck, T.; Strasser, R.; Dommes, J.; Gaspar, T. Hyperhydricity of micropropagated shoots: A typically stress-inducedchange of physiological state plant Cell. Tissue Organ Cult. 2004, 77, 181–191. [Google Scholar] [CrossRef]
- Lu, C.-Y. The use of thidiazuron in tissue culture. Vitr. Cell. Dev. Boil. Anim. 1993, 29, 92–96. [Google Scholar] [CrossRef]
- Kanwar, J.; Kumar, S. Influence of growth regulators and explants on shoot regeneration in carnation. Hortic. Sci. 2009, 36, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Nontaswatsri, C.; Fukai, S.; Touma, T.; Goi, M. Comparison of adventitious shoot formation from node and leaf explants of various carnation (Dianthus caryophyllus L.) cultivars. J. Hortic. Sci. Biotechnol. 2002, 77, 520–525. [Google Scholar] [CrossRef]
- Roberts, A.V.; Smith, E.F.; Mottley, J. The preparation of micropropagated plantlets for transfer to soil without acclimatization. In Methods in Molecular; Pollard, J.W., Walker, J.M., Eds.; Plant Cell and Tissue Culture; Humana Press: Clifton, NJ, USA, 1990; Volume 6, pp. 227–236. [Google Scholar]
- Naing, A.H.; Kim, S.H.; Chung, M.Y.; Kim, C.K. In vitro propagation method for production of morphologically and genetically stable plants of diferent strawberry cultivars. Plant Methods 2019, 15, 36. [Google Scholar] [CrossRef]
- Manikandan, A.; Rachana, D. Response of Carnation to Potassium Fertilizer Sources and Levels Under Polyhouse Conditions. Acta Sci. Agric. 2018, 2, 77–81. [Google Scholar]
- Pe, P.P.W.; Naing, A.H.; Soe, M.T.; Kang, H.H.; Park, K.I.; Kim, C.K. Establishment of meristem culture for virus-free and genetically stable production of the endangered plant Hosta capitata. Sci. Hortic. 2020, 272, 109591. [Google Scholar] [CrossRef]
- Aalifar, M.; Arab, M.; Aliniaeifard, S.; Dianati, S.; Mehrjerdi, M.Z.; Limpens, E.; Serek, M. Embryogenesis efficiency and genetic stability of Dianthus caryophyllus embryos in response to different light spectra and plant growth regulators. Plant Cell Tissue Organ Cult. 2019, 139, 479–492. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.; Miller, R.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Plant Growth Regulators (mg/L) | Shoot Induction (%) * | Mean Number of Shoots/Explants | |
|---|---|---|---|---|
| IBA | TDZ | |||
| 0 | 0 | 0.00 f | 0.00 e | |
| Jinju | 0.1 | 0.2 | 66.67 a | 3.46 a |
| 0.3 | 0.2 | 6.67 e | 4.33 a | |
| 0.5 | 0.2 | 13.33 e | 1.67 c | |
| 1.0 | 0.2 | 16.67de | 1.83 c | |
| 2.0 | 0.2 | 10.00 e | 1.33 c | |
| 0.5 | 0.1 | 53.33 b | 1.40 c | |
| 0.5 | 0.3 | 20.00 cd | 3.89 a | |
| 0.5 | 0.5 | 26.67 cd | 2.53 b | |
| 0.5 | 1.0 | 30.00 c | 0.64 d | |
| 0.5 | 2.0 | 20.00 cd | 3.33 ab | |
| DNV | 0 | 0 | 0.00 d | 0.00 e |
| 0.1 | 0.2 | 10.00 bc | 3.33 ab | |
| 0.3 | 0.2 | 6.67 bc | 3.33 ab | |
| 0.5 | 0.2 | 3.33 c | 0.67 d | |
| 1.0 | 0.2 | 16.67 bc | 3.83 a | |
| 2.0 | 0.2 | 13.33 bc | 1.50 c | |
| 0.5 | 0.1 | 10.00 bc | 1.67 c | |
| 0.5 | 0.3 | 26.67 a | 2.83 b | |
| 0.5 | 0.5 | 16.67 bc | 1.67 c | |
| 0.5 | 1.0 | 16.67 bc | 2.25 b | |
| 0.5 | 2.0 | 13.33 bc | 1.67 c | |
| KB | 0 | 0 | 0.00 d | 0.00 d |
| 0.1 | 0.2 | 26.67 a | 2.90 b | |
| 0.3 | 0.2 | 3.33 c | 0.67 c | |
| 0.5 | 0.2 | 6.67 bc | 0.67 c | |
| 1.0 | 0.2 | 3.33 c | 0.67 c | |
| 2.0 | 0.2 | 3.33 c | 0.67 c | |
| 0.5 | 0.1 | 13.33 bc | 5.00 a | |
| 0.5 | 0.3 | 3.33 c | 0.33 d | |
| 0.5 | 0.5 | 3.33 c | 2.00 b | |
| 0.5 | 1.0 | 6.67 b | 0.67 c | |
| 0.5 | 2.0 | 26.67 a | 0.50 c | |
| Name of Medium | Components |
|---|---|
| Type 1 | MS basal medium [34], 3% sucrose, 0.8% plant agar, pH 5.8 |
| Type 2 | MS Gamborg B5 vitamins medium [35], 2-morpholinoethanesulfonic acid(MES) monohydrate 590 mg/L, BA 0.5 mg/L, NAA 0.02 mg/L, 3% sucrose, 0.25% gelrite, pH 5.8 |
| Type 3 | MS medium, adenine 80 mg/L, sodium phosphate monobasic (NaH2PO4) 85 mg/L, 2% sucrose, 0.8% plant agar, pH 5.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thu, H.T.M.; Naing, A.H.; Jeong, H.Y.; Kim, C.K. Regeneration of Genetically Stable Plants from in Vitro Vitrified Leaves of Different Carnation Cultivars. Plants 2020, 9, 950. https://doi.org/10.3390/plants9080950
Thu HTM, Naing AH, Jeong HY, Kim CK. Regeneration of Genetically Stable Plants from in Vitro Vitrified Leaves of Different Carnation Cultivars. Plants. 2020; 9(8):950. https://doi.org/10.3390/plants9080950
Chicago/Turabian StyleThu, Ho Thi Minh, Aung Htay Naing, Hui Yeong Jeong, and Chang Kil Kim. 2020. "Regeneration of Genetically Stable Plants from in Vitro Vitrified Leaves of Different Carnation Cultivars" Plants 9, no. 8: 950. https://doi.org/10.3390/plants9080950
APA StyleThu, H. T. M., Naing, A. H., Jeong, H. Y., & Kim, C. K. (2020). Regeneration of Genetically Stable Plants from in Vitro Vitrified Leaves of Different Carnation Cultivars. Plants, 9(8), 950. https://doi.org/10.3390/plants9080950