
Evolutionary Game of Vaccination Considering Both Epidemic and Economic Factors by Infectious Network of Complex Nodes

Abstract
:1. Introduction
- (1)
- A novel kind of complex network, including the environment node and aerosol environmental transmission, is presented to consider clustering’s effectiveness during the epidemic process.
- (2)
- Based on the mean-field approximation theory, the endemic equilibrium point and its asymptotic stability in the proposed model under random immunization are demonstrated by making use of a Jacobian matrix and the Lyapunov function to theoretically illustrate the impact of epidemiological characteristics and vaccine features on transmission trends.
- (3)
- The model also combines the topology of the complex network with factors affecting the immunization decision to calculate the number of vaccinated individuals in order to assess the competition of two vaccines under evolutionary game theory, which includes the epidemic risk, information transmission in the neighborhood of the network, and economic factors.
- (4)
- Numerical simulations provide further confirmation of the theoretical results. In the context of vaccines’ market competition, compared with a random immunization strategy, the focus is on how the cost and efficiency of vaccines affect the game between individuals, and thereby affect the prevalence of epidemics.
2. Background
2.1. Epidemiological Models on Complex Networks
- (1)
- The initial network contains isolated nodes. Each time a new node is added into the network, it is connected to m existing nodes ().
- (2)
- The probability of a connection between the new node and the node for which the degree value is can be described as:where is the total degree of the existing nodes in the scale-free network.
- (1)
- Average degree
- (2)
- Average shortest path length
- (3)
- Average clustering coefficient
- (4)
- Diameter
2.2. Game Theory in Infectious Networks
3. Epidemic Model
3.1. Epidemic Transmission Network
3.2. Model under Random Immunization Strategy
3.3. Vaccination Evolutionary Game Model
4. Simulations and Discussion
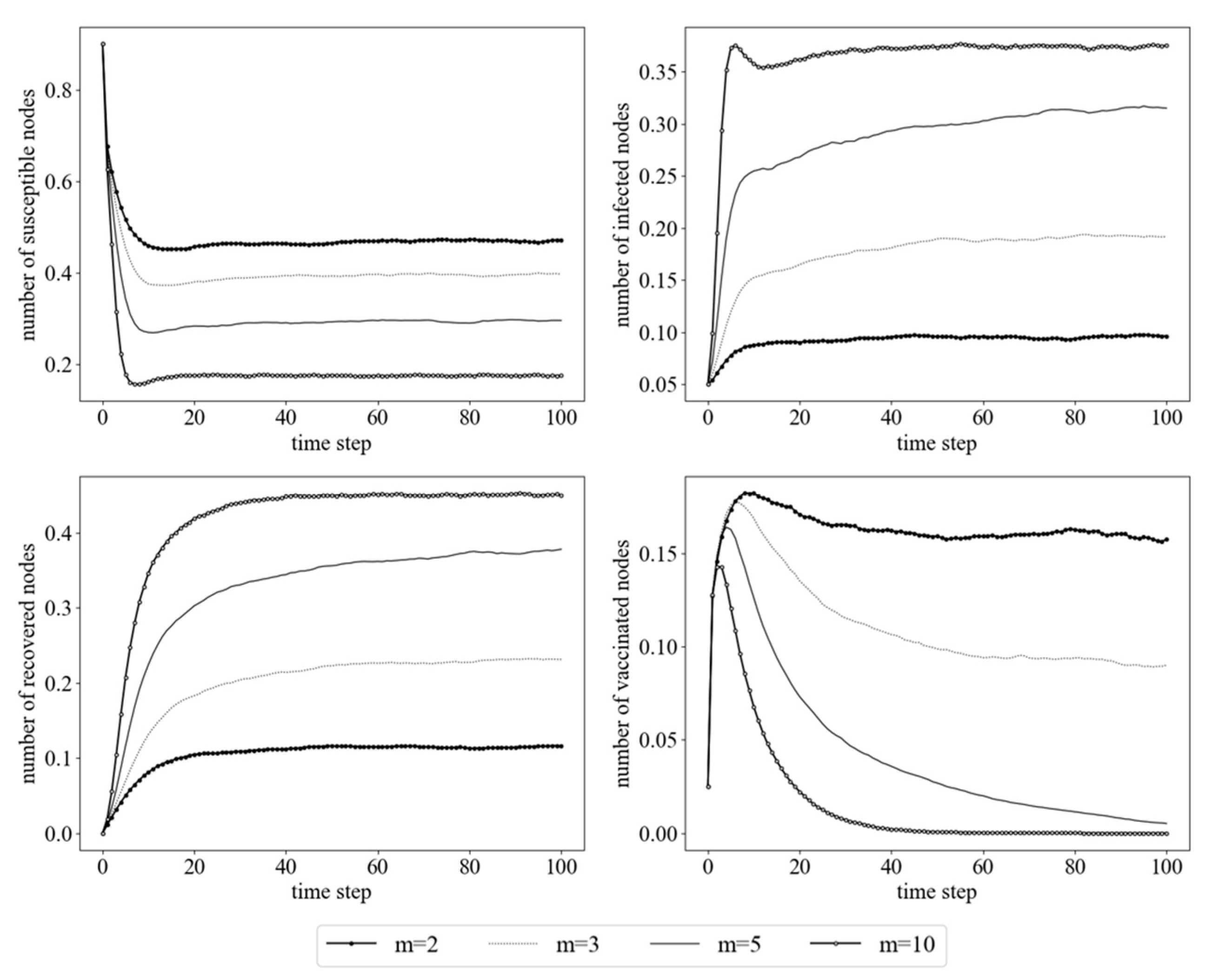
4.1. Simulations of Random Immunization Strategy
4.2. Simulations on Vaccination Evolutionary Game Model
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Elaiw, A.M.; Al Agha, A.D. Global Stability of a Reaction–Diffusion Malaria/COVID-19 Coinfection Dynamics Model. Mathematics 2022, 10, 4390. [Google Scholar] [CrossRef]
- Ramos, A.; Schimit, P. Disease spreading on populations structured by groups. Appl. Math. Comput. 2019, 353, 265–273. [Google Scholar] [CrossRef]
- Wongvanich, N.; Tang, I.; Dubois, M.; Pongsumpun, P. Mathematical Modeling and Optimal Control of the Hand Foot Mouth Disease Affected by Regional Residency in Thailand. Mathematics 2021, 9, 2863. [Google Scholar] [CrossRef]
- Tao, L.; Wang, Y.; Guan, Z.H. Spreading dynamics of a SIQRS epidemic model on scale-free networks. Commun. Nonlinear Sci. Numer. Simul. 2013, 19, 686–692. [Google Scholar]
- Veliov, V.M.; Widder, A. Aggregation and asymptotic analysis of an SI-epidemic model for heterogeneous populations. Math. Med. Biol. J. IMA 2016, 33, 295–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Castillo-Chavez, C. Dynamics of SI models with both horizontal and vertical transmissions as well as Allee effects. Math. Biosci. 2013, 248, 97–116. [Google Scholar] [CrossRef] [Green Version]
- Denes, A.; Muroya, Y.; Rost, G. Global stability of a multistrain SIS model with superinfection. Math. Biosci. Eng. 2016, 14, 421–435. [Google Scholar] [CrossRef]
- Seno, H. An SIS model for the epidemic dynamics with two phases of the human day-to-day activity. J. Math. Biol. 2020, 80, 2109–2140. [Google Scholar] [CrossRef] [Green Version]
- Krause, A.L.; Kurowski, L.; Yawar, K.; Van Gorder, R.A. Stochastic epidemic metapopulation models on networks: SIS dynamics and control strategies. J. Theor. Biol. 2018, 14, 35–52. [Google Scholar] [CrossRef]
- Ayoob, S.; Toktam, M.; Hevi, S.G.; Noreen, I.A.; Ebadi, M.J. SIR model for the spread of COVID-19: A case study. Oper. Res. Perspect. 2023, 10, 100265. [Google Scholar]
- Buonomo, B.; d’Onofrio, A.; Lacitignola, D. Global stability of an SIR epidemic model with information dependent vaccination. Math. Biosci. 2008, 216, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Wickramaarachchi, W.; Perera, S. An SIER model to estimate optimal transmission rate and initial parameters of COVD-19 dynamic in Sri Lanka. Alex. Eng. J. 2020, 60, 1557–1563. [Google Scholar] [CrossRef]
- Trawicki, M.B. Deterministic SEIRS Epidemic Model for Modeling Vital Dynamics, Vaccinations, and Temporary Immunity. Mathmatics 2017, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Blavatska, V.; Holovatch, Y. Spreading processes in “post-epidemic” environments. II. Safety patterns on scale-free networks. Phys. A Stat. Mech. Its Appl. 2022, 591, 126799. [Google Scholar] [CrossRef]
- Bing, W.; Hongjuan, Z.; Yuexing, H. Dynamical immunization based on random-walk in time-varying networks. Chaos Solitons Fractals 2022, 155, 111755. [Google Scholar]
- Bai, W.H.; Zhou, T.; Wang, B.H. Immunization of susceptible-infected model on scale-free networks. Phys. A Stat. Mech. Appl. 2007, 384, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Lihong, S.; Qiang, H.; Yueyang, T.; Qi, Z.; Xin, Y.; Xingwei, W. A complex network-based vaccination strategy for infectious diseases. Appl. Soft Comput. 2023, 136, 110081. [Google Scholar]
- Madar, N.; Kalisky, T.; Cohen, R.; Ben-Avraham, D.; Havlin, S. Immunization and epidemic dynamics in complex networks. Eur. Phys. J. B 2004, 38, 269–276. [Google Scholar] [CrossRef]
- Masaki, T.; Jun, T. Proposal of an apposite strategy-updating rule for the vaccination game where hubs refer to hubs and lower-degree agents refer to lower-degree agents. Biosystems 2021, 209, 104532. [Google Scholar]
- Liu, Y.; Jian, S.; Gao, J. Dynamics analysis and optimal control of SIVR epidemic model with incomplete immunity. Adv. Contin. Discret. Model. 2022, 2022, 51. [Google Scholar] [CrossRef]
- Buonomo, B.; Lacitignola, D.; Vargas-De-Leon, C. Qualitative analysis and optimal control of an epidemic model with vaccination and treatment. Math. Comput. Simul. 2014, 100, 88–102. [Google Scholar] [CrossRef]
- Angran, L.; Faisal, Y.; Zeeshan, A.; Waqas, N. Analytical solution of a non-linear fractional order SIS epidemic model utilizing a new technique. Alex. Eng. J. 2023, 73, 123–129. [Google Scholar]
- Liu, X.N.; Takeuchi, Y.; Iwami, S. SVIR epidemic models with vaccination strategies. J. Theor. Biol. 2008, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Haus, B.; Mercorelli, P. Extension of SEIR Compartmental Models for Constructive Lyapunov Control of COVID-19 and Analysis in Terms of Practical Stability. Mathematics 2021, 9, 2076. [Google Scholar] [CrossRef]
- Xiao, Y.; Tang, S. Dynamics of infection with nonlinear incidence in a simple vaccination model. Nonlinear Anal. Real World Appl. 2010, 11, 4154–4163. [Google Scholar] [CrossRef]
- Mirzasoleiman, B.; Babaei, M.; Jalili, M. Immunizing complex networks with limited budget. EPL 2012, 98, 38004. [Google Scholar] [CrossRef]
- Duan, X.C.; Yuan, S.L.; Li, X.Z. Global stability of an SVIR model with age of vaccination. Appl. Math. Comput. 2014, 226, 528–540. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, P.K.; Dong, Y.; Takeuchi, Y. Optimal control of infectious disease: Information-induced vaccination and limited treatment. Phys. A Stat. Mech. Its Appl. 2020, 542, 123196. [Google Scholar] [CrossRef]
- Arefin, M.R.; Masaki, T.; Tanimoto, J. Vaccinating behaviour guided by imitation and aspiration. Proceedings of the Royal Society. Math. Phys. Eng. Sci. 2020, 476, 20200327. [Google Scholar]
- Liu, J.; Zhang, T. Epidemic spreading of an SEIRS model in scale-free networks. Commun. Nonlinear Sci. Numer. Simul. 2011, 16, 3375–3384. [Google Scholar] [CrossRef]
- WHO. Ten Threats to Global Health in 2019. Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (accessed on 11 January 2023).
- Nuwarda, R.F.; Ramzan, I.; Weekes, L.; Kayser, V. Vaccine Hesitancy: Contemporary Issues and Historical Background. Vaccines 2022, 10, 1595. [Google Scholar] [CrossRef] [PubMed]
- Fine, P.; Eames, K.; Heymann, D.L. “Herd Immunity’’: A Rough Guide. Clin. Infect. Dis. 2011, 52, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Streefland, P.H. Public doubts about vaccination safety and resistance against vaccination. Health Policy 2001, 55, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Cardillo, A.; Reyes-Suarez, C.; Naranjo, F.; Gomez-Gardenes, J. Evolutionary vaccination dilemma in complex networks. Phys. Rev. E 2013, 88, 32803. [Google Scholar] [CrossRef] [Green Version]
- Bauch, C.T. Imitation dynamics predict vaccinating behaviour. Proc. R. Soc. B Biol. Sci. 2005, 272, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.; Kabir, K.M.A.; Tanimoto, J. Based on mathematical epidemiology and evolutionary game theory, which is more effective: Quarantine or isolation policy? J. Stat. Mech. Theory Exp. 2020, 3, 033502. [Google Scholar] [CrossRef]
- Hauert, C.; Doebeli, M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature 2004, 428, 643–646. [Google Scholar] [CrossRef]
- Zhong, S.; Wu, X.Q.; Li, Y.T.; Liu, C.Y. Indirect transmission and disinfection strategies on heterogeneous networks. Phys. Rev. E 2022, 106, 054309. [Google Scholar] [CrossRef]
- Granell, C.; Mucha, P.J. Epidemic spreading in localized environments with recurrent mobility patterns. Phys. Rev. E 2018, 97, 052302. [Google Scholar] [CrossRef] [Green Version]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Topological Property | m = 2 | m = 3 | m = 5 | m = 10 |
|---|---|---|---|---|---|
| 3000 | average degree | 3.997 | 5.994 | 9.983 | 19.933 |
| average shortest path length | 4.557 | 3.864 | 3.316 | 2.805 | |
| average clustering | 0.012 | 0.014 | 0.018 | 0.028 | |
| diameter | 8 | 6 | 5 | 4 | |
| 5000 | average degree | 3.998 | 5.996 | 9.990 | 19.960 |
| average shortest path length | 4.775 | 4.051 | 3.469 | 2.910 | |
| average clustering | 0.007 | 0.009 | 0.012 | 0.019 | |
| diameter | 8 | 7 | 5 | 4 | |
| 10,000 | average degree | 3.999 | 5.998 | 9.995 | 19.980 |
| average shortest path length | 5.047 | 4.307 | 3.670 | 3.062 | |
| average clustering | 0.003 | 0.005 | 0.007 | 0.011 | |
| diameter | 9 | 7 | 6 | 5 | |
| 20,000 | average degree | 4.000 | 5.999 | 9.998 | 19.990 |
| average shortest path length | 5.316 | 4.522 | 3.859 | 3.237 | |
| average clustering | 0.003 | 0.003 | 0.004 | 0.007 | |
| diameter | 9 | 8 | 6 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Xiang, Z. Evolutionary Game of Vaccination Considering Both Epidemic and Economic Factors by Infectious Network of Complex Nodes. Mathematics 2023, 11, 2697. https://doi.org/10.3390/math11122697
Li B, Xiang Z. Evolutionary Game of Vaccination Considering Both Epidemic and Economic Factors by Infectious Network of Complex Nodes. Mathematics. 2023; 11(12):2697. https://doi.org/10.3390/math11122697
Chicago/Turabian StyleLi, Bing, and Ziye Xiang. 2023. "Evolutionary Game of Vaccination Considering Both Epidemic and Economic Factors by Infectious Network of Complex Nodes" Mathematics 11, no. 12: 2697. https://doi.org/10.3390/math11122697