
Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of Papain-like Cysteine Proteases Primary Structure
2.2. Specific IgE Detection with the FABER® Multiplex Testing System
2.3. Patients
3. Results
3.1. Comparison of Primary Structures
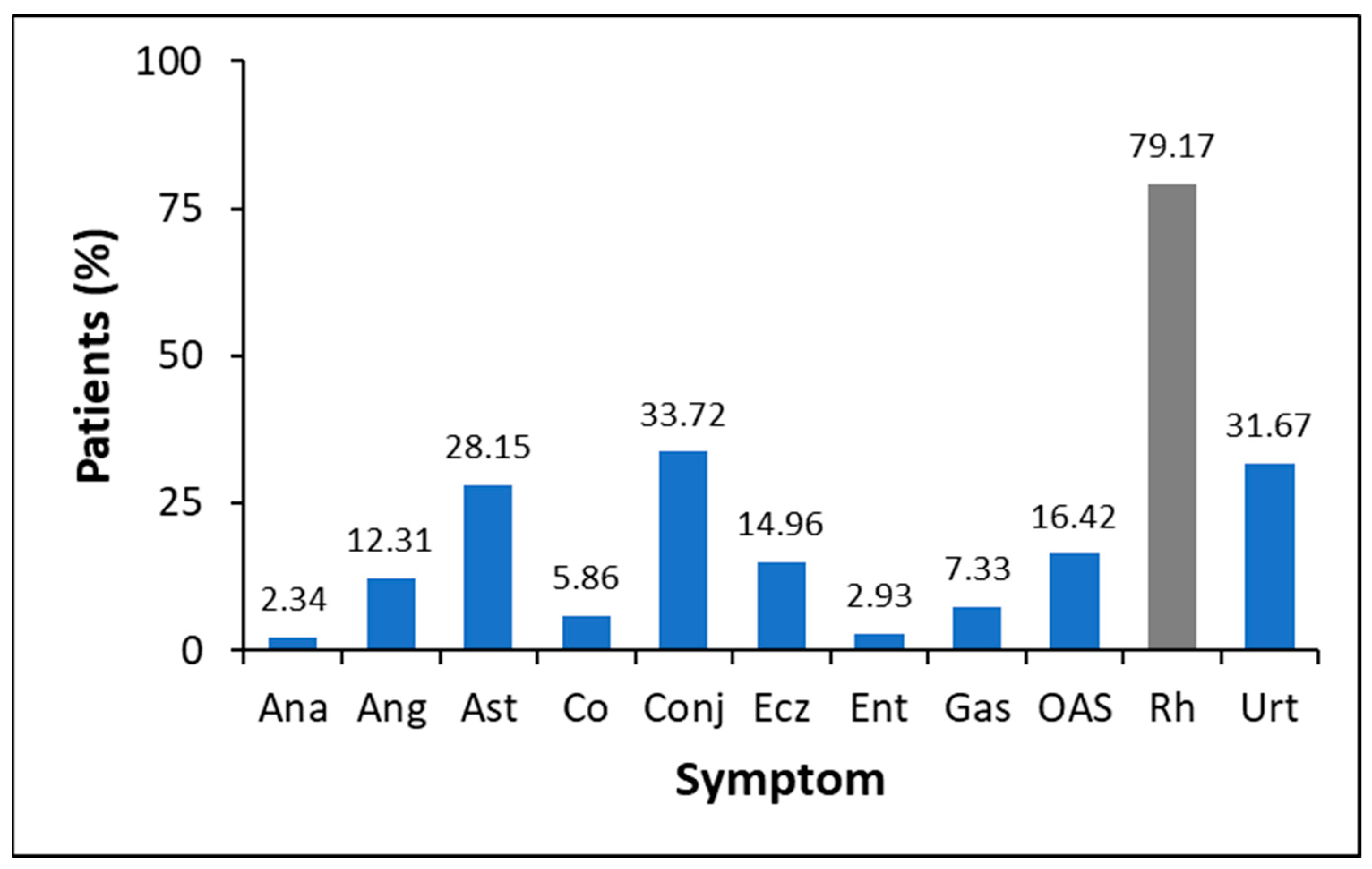
3.2. Allergy Symptoms of the Analysed Population
3.3. Results of Specific IgE Detection by In Vitro Tests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petushkova, A.I.; Savvateeva, L.V.; Zamyatnin, A.A. Structure determinants defining the specificity of papain-like cysteine proteases. Comput. Struct. Biotechnol. J. 2022, 20, 6552–6569. [Google Scholar] [CrossRef]
- Wu, W.U.; Hettiarachchy, N.S.; Qi, M. Hydrophobicity, solubility, and emulsifying properties of soy protein peptides prepared by papain modification and ultrafiltration. J. Am. Oil Chem. Soc. 1998, 75, 845–850. [Google Scholar] [CrossRef]
- Polaina, J.; MacCabe, A.P. Industrial Enzymes; Springer: Dordrecht, The Netherlands, 2007; pp. 181–195. Available online: https://link.springer.com/book/10.1007/1-4020-5377-0 (accessed on 15 March 2023).
- Verma, S.; Dixit, R.; Pandey, K.C. Cysteine Proteases: Modes of Activation and Future Prospects as Pharmacological Targets. Front. Pharmacol. 2016, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Lynn, K.R.; Yaguchi, M. N-Terminal homology in three cysteinyl proteases from Papaya latex. Biochim. Biophys. Acta 1979, 581, 363–364. [Google Scholar] [CrossRef]
- Yamada, F.; Takahashi, N.; Murachi, T. Purification and characterization of a proteinase from pineapple fruit, fruit bromelain FA2. J. Biochem. 1976, 79, 1223–1234. [Google Scholar] [CrossRef]
- Matagne, A.; Bolle, L.; El Mahyaoui, R.; Baeyens-Volant, D.; Azarkan, M. The proteolytic system of pineapple stems revisited: Purification and characterization of multiple catalytically active forms. Phytochemistry 2017, 138, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Pastorello, E.A.; Conti, A.; Pravettoni, V.; Farioli, L.; Rivolta, F.; Ansaloni, R.; Ispano, M.; Incorvaia, C.; Giuffrida, M.G.; Ortolani, C. Identification of actinidin as the major allergen of kiwi fruit. J. Allergy Clin. Immunol. 1998, 101, 531–537. [Google Scholar] [CrossRef]
- Tuppo, L.; Giangrieco, I.; Palazzo, P.; Bernardi, M.L.; Scala, E.; Carratore, V.; Tamburrini, M.; Mari, A.; Ciardiello, M.A. Kiwellin, a modular protein from green and gold kiwi fruits: Evidence of in vivo and in vitro processing and IgE binding. J. Agric. Food Chem. 2008, 56, 3812–3817. [Google Scholar] [CrossRef]
- Misas-Villamil, J.C.; van der Hoorn, R.A.; Doehlemann, G. Papain-like cysteine proteases as hubs in plant immunity. New Phytol. 2016, 212, 902–907. [Google Scholar] [CrossRef]
- Jiang, N.; Yin, J.; Wen, L. Papain Induced Occupational Asthma with Kiwi and Fig Allergy. Allergy Asthma Immunol. Res. 2016, 8, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Drenth, J.; Jansonius, J.N.; Koekoek, R.; Swen, H.M.; Wolthers, B.G. Structure of papain. Nature 1968, 218, 929–932. [Google Scholar] [CrossRef]
- Vernet, T.; Tessier, D.C.; Chatellier, J.; Plouffe, C.; Lee, T.S.; Thomas, D.Y.; Storer, A.C.; Ménard, R. Structural and functional roles of asparagine 175 in the cysteine protease papain. J. Biol. Chem. 1995, 270, 16645–16652. [Google Scholar] [CrossRef] [PubMed]
- Novinec, M. Computational investigation of conformational variability and allostery in cathepsin K and other related peptidases. PLoS ONE 2017, 12, e0182387. [Google Scholar] [CrossRef]
- Kamphuis, I.G.; Drenth, J.; Baker, E.N. Thiol proteases. Comparative studies based on the high-resolution structures of papain and actinidin, and on amino acid sequence information for cathepsins B and H, and stem bromelain. J. Mol. Biol. 1985, 182, 317–329. [Google Scholar] [CrossRef]
- Tanabe, S.; Arai, S.; Watanabe, M. Modification of wheat flour with bromelain and baking hypoallergenic bread with added ingredients. Biosci. Biotechnol. Biochem. 1996, 60, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Freddi, G.; Mossotti, R.; Innocenti, R. Degumming of silk fabric with several proteases. J. Biotechnol. 2003, 106, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Arshad, Z.I.; Amid, A.; Yusof, F.; Jaswir, I.; Ahmad, K.; Loke, S.P. Bromelain: An overview of industrial application and purification strategies. Appl. Microbiol. Biotechnol. 2014, 98, 7283–7297. [Google Scholar] [CrossRef] [PubMed]
- Aminlari, M.; Shekarforoush, S.S.; Gheisari, H.R.; Golestan, L. Effect of actinidin on the protein solubility, water holding capacity, texture, electrophoretic pattern of beef, and on the quality attributes of a sausage product. J. Food Sci. 2009, 74, C221–C226. [Google Scholar] [CrossRef]
- Bekhit, A.A.; Hopkins, D.L.; Geesink, G.; Franks, P. Exogenous proteases for meat tenderization. Crit. Rev. Food Sci. Nutr. 2014, 54, 1012–1031. [Google Scholar] [CrossRef] [PubMed]
- Aider, M. Potential applications of ficin in the production of traditional cheeses and protein hydrolysates. JDS Commun. 2021, 2, 233–237. [Google Scholar] [CrossRef]
- Thomas, W.R.; Stewart, G.A.; Simpson, R.J.; Chua, K.Y.; Plozza, T.M.; Dilworth, R.J.; Nisbet, A.; Turner, K.J. Cloning and expression of DNA coding for the major house dust mite allergen Der p 1 in Escherichia coli. Int. Arch. Allergy Appl. Immunol. 1988, 85, 127–129. [Google Scholar] [CrossRef]
- Mills, E.N.; Jenkins, J.A.; Alcocer, M.J.; Shewry, P.R. Structural, biological, and evolutionary relationships of plant food allergens sensitizing via the gastrointestinal tract. Crit. Rev. Food Sci. Nutr. 2004, 44, 379–407. [Google Scholar] [CrossRef]
- Breiteneder, H.; Radauer, C. A classification of plant food allergens. J. Allergy Clin. Immunol. 2004, 113, 821–830. [Google Scholar] [CrossRef]
- Urbani, S.; Aruanno, A.; Nucera, E. Adverse reaction to. Clin. Mol. Allergy 2020, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Pomés, A.; Davies, J.M.; Gadermaier, G.; Hilger, C.; Holzhauser, T.; Lidholm, J.; Lopata, A.L.; Mueller, G.A.; Nandy, A.; Radauer, C.; et al. WHO/IUIS Allergen Nomenclature: Providing a common language. Mol. Immunol. 2018, 100, 3–13. [Google Scholar] [CrossRef]
- Glesner, J.; Vailes, L.D.; Schlachter, C.; Mank, N.; Minor, W.; Osinski, T.; Chruszcz, M.; Chapman, M.D.; Pomés, A. Antigenic Determinants of Der p 1: Specificity and Cross-Reactivity Associated with IgE Antibody Recognition. J. Immunol. 2017, 198, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Bublin, M.; Dennstedt, S.; Buchegger, M.; Antonietta Ciardiello, M.; Bernardi, M.L.; Tuppo, L.; Harwanegg, C.; Hafner, C.; Ebner, C.; Ballmer-Weber, B.K.; et al. The performance of a component-based allergen microarray for the diagnosis of kiwifruit allergy. Clin. Exp. Allergy 2011, 41, 129–136. [Google Scholar] [CrossRef]
- Gailhofer, G.; Wilders-Truschnig, M.; Smolle, J.; Ludvan, M. Asthma caused by bromelain: An occupational allergy. Clin. Exp. Allergy 1988, 18, 445–450. [Google Scholar] [CrossRef]
- Bhowmik, M.; Biswas Sarkar, M.; Kanti Sarkar, R.; Dasgupta, A.; Saha, S.; Jana, K.; Sircar, G.; Gupta Bhattacharya, S. Cloning and immunobiochemical analyses on recombinant chymopapain allergen Cari p 2 showing pollen-fruit cross-reaction. Mol. Immunol. 2021, 137, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kampen, V.v.; Merget, R.; Brüning, T. Occupational allergies to papain. Pneumologie 2005, 59, 405–410. [Google Scholar] [CrossRef]
- Tymoszuk, D.; Wiszniewska, M.; Walusiak-Skorupa, J. Papain-induced occupational rhinoconjunctivitis and asthma—A case report. Med. Pract. 2016, 67, 109–112. [Google Scholar] [CrossRef]
- Mansfield, L.E.; Bowers, C.H. Systemic reaction to papain in a nonoccupational setting. J. Allergy Clin. Immunol. 1983, 71, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Soh, W.T.; Zhang, J.; Hollenberg, M.D.; Vliagoftis, H.; Rothenberg, M.E.; Sokol, C.L.; Robinson, C.; Jacquet, A. Protease allergens as initiators-regulators of allergic inflammation. Allergy 2023, 78, 1148–1168. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, C.; Ferrara, R.; Bernardi, M.L.; Zennaro, D.; Tuppo, L.; Giangrieco, I.; Ricciardi, T.; Tamburrini, M.; Ciardiello, M.A.; Mari, A. Molecular approach to a patient’s tailored diagnosis of the oral allergy syndrome. Clin. Transl. Allergy 2020, 10, 22. [Google Scholar] [CrossRef]
- Giangrieco, I.; Ricciardi, T.; Alessandri, C.; Farina, L.; Crescenzo, R.; Tuppo, L.; Ciancamerla, M.; Rafaiani, C.; Bernardi, M.L.; Digilio, A.F.; et al. ENEA, a peach and apricot IgE-binding protein cross-reacting with the latex major allergen Hev b 5. Mol. Immunol. 2019, 112, 347–357. [Google Scholar] [CrossRef]
- Ricciardi, T.; Giangrieco, I.; Alessandri, C.; Rafaiani, C.; Tuppo, L.; Tamburrini, M.; Digilio, A.F.; Ciardiello, M.A.; Mari, A. Pattern of sensitization to Juniperus oxycedrus 4EF-hand polcalcin, Jun o 4, compared with the 2EF-hand grass homolog Phl p 7 in a general Italian population of subjects suffering from pollinosis. Clin. Immunol. 2022, 234, 108894. [Google Scholar] [CrossRef]
- Tuppo, L.; Giangrieco, I.; Tamburrini, M.; Alessandri, C.; Mari, A.; Ciardiello, M.A. Detection of Allergenic Proteins in Foodstuffs: Advantages of the Innovative Multiplex Allergen Microarray-Based Immunoassay Compared to Conventional Methods. Foods 2022, 11, 878. [Google Scholar] [CrossRef]
- D’Avino, R.; Bernardi, M.L.; Wallner, M.; Palazzo, P.; Camardella, L.; Tuppo, L.; Alessandri, C.; Breiteneder, H.; Ferreira, F.; Ciardiello, M.A.; et al. Kiwifruit Act d 11 is the first member of the ripening-related protein family identified as an allergen. Allergy 2011, 66, 870–877. [Google Scholar] [CrossRef]
- Al-Ghouleh, A.; Johal, R.; Sharquie, I.K.; Emara, M.; Harrington, H.; Shakib, F.; Ghaemmaghami, A.M. The glycosylation pattern of common allergens: The recognition and uptake of Der p 1 by epithelial and dendritic cells is carbohydrate dependent. PLoS ONE 2012, 7, e33929. [Google Scholar] [CrossRef]
- Deleuran, M.; Ellingsen, A.R.; Paludan, K.; Schou, C.; Thestrup-Pedersen, K. Purified Der p1 and p2 patch tests in patients with atopic dermatitis: Evidence for both allergenicity and proteolytic irritancy. Acta Derm. Venereol. 1998, 78, 241–243. [Google Scholar] [CrossRef]
- Custovic, A.; Taggart, S.C.; Francis, H.C.; Chapman, M.D.; Woodcock, A. Exposure to house dust mite allergens and the clinical activity of asthma. J. Allergy Clin. Immunol. 1996, 98, 64–72. [Google Scholar] [CrossRef]
- Sanchez-Borges, M.; Capriles-Hulett, A.; Fernandez-Caldas, E.; Suarez-Chacon, R.; Caballero, F.; Castillo, S.; Sotillo, E. Mite-contaminated foods as a cause of anaphylaxis. J. Allergy Clin. Immunol. 1997, 99, 738–743. [Google Scholar] [CrossRef]
- van der Heide, S.; De Monchy, J.G.; De Vries, K.; Dubois, A.E.; Kauffman, H.F. Seasonal differences in airway hyperresponsiveness in asthmatic patients: Relationship with allergen exposure and sensitization to house dust mites. Clin. Exp. Allergy 1997, 27, 627–633. [Google Scholar] [CrossRef]
- Ogi, K.; Ramezanpour, M.; Liu, S.; Ferdoush Tuli, J.; Bennett, C.; Suzuki, M.; Fujieda, S.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Der p 1 Disrupts the Epithelial Barrier and Induces IL-6 Production in Patients with House Dust Mite Allergic Rhinitis. Front. Allergy 2021, 2, 692049. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Chapman, M.D.; Vailes, L.D.; Stura, E.A.; Saint-Remy, J.M.; Minor, W.; Pomés, A. Crystal structures of mite allergens Der f 1 and Der p 1 reveal differences in surface-exposed residues that may influence antibody binding. J. Mol. Biol. 2009, 386, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Heymann, P.W.; Chapman, M.D.; Platts-Mills, T.A. Antigen Der f I from the dust mite Dermatophagoides farinae: Structural comparison with Der p I from Dermatophagoides pteronyssinus and epitope specificity of murine IgG and human IgE antibodies. J. Immunol. 1986, 137, 2841–2847. Available online: https://pubmed.ncbi.nlm.nih.gov/2428875/ (accessed on 29 May 2023). [PubMed]
- Jeong, K.Y.; Park, J.W.; Hong, C.S. House dust mite allergy in Korea: The most important inhalant allergen in current and future. Allergy Asthma Immunol. Res. 2012, 4, 313–325. [Google Scholar] [CrossRef]
- Krämer, U.; Lemmen, C.; Bartusel, E.; Link, E.; Ring, J.; Behrendt, H. Current eczema in children is related to Der f 1 exposure but not to Der p 1 exposure. Br. J. Dermatol. 2006, 154, 99–105. [Google Scholar] [CrossRef]
- Lim, F.L.; Hashim, Z.; Than, L.T.; Md Said, S.; Hisham Hashim, J.; Norbäck, D. Asthma, Airway Symptoms and Rhinitis in Office Workers in Malaysia: Associations with House Dust Mite (HDM) Allergy, Cat Allergy and Levels of House Dust Mite Allergens in Office Dust. PLoS ONE 2015, 10, e0124905. [Google Scholar] [CrossRef]
- Azarkan, M.; El Moussaoui, A.; van Wuytswinkel, D.; Dehon, G.; Looze, Y. Fractionation and purification of the enzymes stored in the latex of Carica papaya. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 790, 229–238. [Google Scholar] [CrossRef]
- Drugs and Lactation Database (LactMed®) [Internet]; Papaya; National Institute of Child Health and Human Development: Bethesda, MD, USA, 2006. Available online: https://pubmed.ncbi.nlm.nih.gov/30000941/ (accessed on 15 April 2023).
- Novey, H.S.; Keenan, W.J.; Fairshter, R.D.; Wells, I.D.; Wilson, A.F.; Culver, B.D. Pulmonary disease in workers exposed to papain: Clinico-physiological and immunological studies. Clin. Exp. Allergy 1980, 10, 721–731. [Google Scholar] [CrossRef]
- Mansfield, L.E.; Ting, S.; Haverly, R.W.; Yoo, T.J. The incidence and clinical implications of hypersensitivity to papain in an allergic population, confirmed by blinded oral challenge. Ann. Allergy 1985, 55, 541–543. Available online: https://pubmed.ncbi.nlm.nih.gov/4051260/ (accessed on 7 July 2023). [PubMed]
- Freye, H.B. Papain anaphylaxis: A case report. Allergy Proc. 1988, 9, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Grammer, L.C.; Patterson, R. Proteins: Chymopapain and insulin. J. Allergy Clin. Immunol. 1984, 74, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Pinkowski, J.L.; Leeson, M.C. Anaphylactic shock associated with chymopapain skin test. A case report and review of the literature. Clin. Orthop. Relat. Res. 1990, 260, 186–190. Available online: https://journals.lww.com/clinorthop/Abstract/1990/11000/Anaphylactic_Shock_Associated_With_Chymopapain.30.aspx (accessed on 21 May 2023).
- Azarkan, M.; Maquoi, E.; Delbrassine, F.; Herman, R.; M’Rabet, N.; Calvo Esposito, R.; Charlier, P.; Kerff, F. Structures of the free and inhibitors-bound forms of bromelain and ananain from Ananas comosus stem and in vitro study of their cytotoxicity. Sci. Rep. 2020, 10, 19570. [Google Scholar] [CrossRef]
- Lida, S.; Sasaki, M.; Ota, S. Immunological cross-reaction between thiol proteases of plant origin: Stem and fruit bromelains. J. Biochem. 1973, 73, 377–386. Available online: https://pubmed.ncbi.nlm.nih.gov/4196855/ (accessed on 21 May 2023).
- Baur, X.; Fruhmann, G. Allergic reactions, including asthma, to the pineapple protease bromelain following occupational exposure. Clin. Exp. Allergy 1979, 9, 443–450. [Google Scholar] [CrossRef]
- Ciardiello, M.A.; Giangrieco, I.; Tuppo, L.; Tamburrini, M.; Buccheri, M.; Palazzo, P.; Bernardi, M.L.; Ferrara, R.; Mari, A. Influence of the natural ripening stage, cold storage, and ethylene treatment on the protein and IgE-binding profiles of green and gold kiwi fruit extracts. J. Agric. Food Chem. 2009, 57, 1565–1571. [Google Scholar] [CrossRef]
- Grozdanovic, M.; Popovic, M.; Polovic, N.; Burazer, L.; Vuckovic, O.; Atanaskovic-Markovic, M.; Lindner, B.; Petersen, A.; Gavrovic-Jankulovic, M. Evaluation of IgE reactivity of active and thermally inactivated actinidin, a biomarker of kiwifruit allergy. Food Chem. Toxicol. 2012, 50, 1013–1018. [Google Scholar] [CrossRef]
- Palacin, A.; Rodriguez, J.; Blanco, C.; Lopez-Torrejon, G.; Sánchez-Monge, R.; Varela, J.; Jiménez, M.A.; Cumplido, J.; Carrillo, T.; Crespo, J.F.; et al. Immunoglobulin E recognition patterns to purified Kiwifruit (Actinidinia deliciosa) allergens in patients sensitized to Kiwi with different clinical symptoms. Clin. Exp. Allergy 2008, 38, 1220–1228. [Google Scholar] [CrossRef]
- Haesaerts, S.; Rodriguez Buitrago, J.A.; Loris, R.; Baeyens-Volant, D.; Azarkan, M. Crystallization and preliminary X-ray analysis of four cysteine proteases from Ficus carica latex. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Raskovic, B.; Lazic, J.; Polovic, N. Characterisation of general proteolytic, milk clotting and antifungal activity of Ficus carica latex during fruit ripening. J. Sci. Food Agric. 2016, 96, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Siar, E.H.; Morellon-Sterling, R.; Zidoune, M.N.; Fernandez-Lafuente, R. Use of glyoxyl-agarose immobilized ficin extract in milk coagulation: Unexpected importance of the ficin loading on the biocatalysts. Int. J. Biol. Macromol. 2020, 144, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Mari, A.; Iacovacci, P.; Afferni, C.; Barletta, B.; Tinghino, R.; Di Felice, G.; Pini, C. Specific IgE to cross-reactive carbohydrate determinants strongly affect the in vitro diagnosis of allergic diseases. J. Allergy Clin. Immunol. 1999, 103, 1005–1011. [Google Scholar] [CrossRef]
- Coutinho, V.; Vidal, C.; Garrido, M.; Gude, F.; Lojo, S.; Linneberg, A.; Gonzalez-Quintela, A. Interference of cross-reactive carbohydrates in the determination of specific IgE in alcohol drinkers and strategies to minimize it: The example of latex. Ann. Allergy Asthma Immunol. 2008, 101, 394–401. [Google Scholar] [CrossRef]
- Abram, D.M.; Fernandes, L.G.R.; Perez-Riverol, A.; Regina Brochetto-Braga, M.; Zollner, R.L. Cross-Reactive Carbohydrate Determinant in. Toxins 2020, 12, 649. [Google Scholar] [CrossRef]
- Chapman, M.D.; Heymann, P.W.; Platts-Mills, T.A. Epitope mapping of two major inhalant allergens, Der p I and Der f I, from mites of the genus Dermatophagoides. J. Immunol. 1987, 139, 1479–1484. [Google Scholar]
- Arthur, R.P.; Shelley, W.B. The role of proteolytic enzymes in the production of pruritus in man. J. Investig. Dermatol. 1955, 25, 341–346. [Google Scholar] [CrossRef]
- Malewicz, N.M.; Ieong, H.F.; Oeck, S.; Kumowski, N.; Zhang, Z.; Shimada, S.G.; LaMotte, R.H. Punctate Administration of Ficin as a human and animal model of non-histaminergic itch. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1300–1307. [Google Scholar] [CrossRef]
- Alessandri, C.; Giangrieco, I.; Tuppo, L.; Ferrara, R.; Zennaro, D.; Bernardi, M.L.; Ciancamerla, M.; Rafaiani, C.; Tamburrini, M.; Mari, A.; et al. Are peas a safe food for lipid transfer protein allergic patients? Allergy 2021, 76, 2587–2589. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Protein Name | N-glycosylation Sites a | Allergen Name | Organism (Scientific Name) | Source Tissue | Route of Exposure | Who Is Exposed | Reported Symptoms b Include |
|---|---|---|---|---|---|---|---|
| Der p 1 | Yes, one [40] | Der p 1 | Dermatophagoides pteronyssinus | Whole body [41] | Inhalation [42] Ingestion [43] | General population [41] | Respiratory symptoms, asthma breathlessness, angioedema, wheezing, rhinorrhea, anaphylaxis [43,44,45] |
| Der f 1 | Yes, one [46] | Der f 1 | Dermatophagoides farinae | Whole body [47] | Inhalation, [48], Ingestion [43] | General population [49] | Respiratory symptoms, asthma breathlessness, angioedema, wheezing, rhinorrhea, anaphylaxis [43,44,50] |
| Papain | Not found in the analysed isoforms | Cari p Papain | Carica papaya | latex, unripe fruit [51,52] | Inhalation, skin contact, [32,53] Ingestion [33,54] | General population, occupational [32] | Respiratory symptoms, rhinoconjunctivitis, asthma anaphylaxis [30,31,55] |
| Chymopapain | Not found in the analysed isoforms | Cari p 2 | Carica papaya | Latex, unripe fruit [52] | Inhalation, ingestion [28,56] | General population, occupational [56] | Respiratory, gastrointestinal, anaphylaxis [28,57] |
| Bromelain | Yes, one (some isoforms only) [58] | Ana c 2 | Ananas comosus | Fruit, stem [59] | Ingestion, inhalation [27] | General population, occupational [27] | gastrointestinal symptoms, asthma [60] |
| Actinidin | Not found in the analysed isoforms | Act d 1 | Actinidia deliciosa | fruit [61] | Ingestion [62] | General population [63] | OAS, respiratory and gastrointestinal symptoms, anaphylaxis [8,26] |
| Ficin | Not found in the analysed isoforms | Fic c Ficin | Ficus carica | Latex, fruit (highest amount in unripe fruit [64,65] | Ingestion, occupational | General population [66], occupational |
| Der p 1 | Der f 1 | Ana c 2 | Cari p Papain | Cari p 2 | Act d 1 | Fic c Ficin | |
|---|---|---|---|---|---|---|---|
| Der p 1 | 100 | ||||||
| Der f 1 | 82.43 | 100 | |||||
| Ana c 2 | 29.06 | 30.05 | 100 | ||||
| Cari p Papain | 29.21 | 30.20 | 41.15 | 100 | |||
| Cari p 2 | 32.21 | 32.69 | 51.43 | 59.43 | 100 | ||
| Act d 1 | 34.13 | 36.06 | 47.14 | 49.29 | 51.15 | 100 | |
| Fic c Ficin | 38.35 | 37.86 | 50.72 | 50.95 | 51.87 | 56.94 | 100 |
| Der p 1 | Der f 1 | Ana c 2 | Cari p Papain | Cari p 2 | Act d 1 | Fic c Ficin | |
|---|---|---|---|---|---|---|---|
| Der p 1 | 67 | ||||||
| Der f 1 | 58 | 73 | |||||
| Ana c 2 | 10 | 10 | 41 | ||||
| Cari p Papain | 2 | 2 | 3 | 5 | |||
| Cari p 2 | 3 | 2 | 5 | 2 | 9 | ||
| Act d 1 | 0 | 0 | 3 | 1 | 0 | 3 | |
| Fic c Ficin | 6 | 6 | 4 | 0 | 0 | 0 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giangrieco, I.; Ciardiello, M.A.; Tamburrini, M.; Tuppo, L.; Rafaiani, C.; Mari, A.; Alessandri, C. Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population. Foods 2023, 12, 2852. https://doi.org/10.3390/foods12152852
Giangrieco I, Ciardiello MA, Tamburrini M, Tuppo L, Rafaiani C, Mari A, Alessandri C. Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population. Foods. 2023; 12(15):2852. https://doi.org/10.3390/foods12152852
Chicago/Turabian StyleGiangrieco, Ivana, Maria Antonietta Ciardiello, Maurizio Tamburrini, Lisa Tuppo, Chiara Rafaiani, Adriano Mari, and Claudia Alessandri. 2023. "Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population" Foods 12, no. 15: 2852. https://doi.org/10.3390/foods12152852
APA StyleGiangrieco, I., Ciardiello, M. A., Tamburrini, M., Tuppo, L., Rafaiani, C., Mari, A., & Alessandri, C. (2023). Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population. Foods, 12(15), 2852. https://doi.org/10.3390/foods12152852