Compatibility of Aerial and Terrestrial LiDAR for Quantifying Forest Structural Diversity

 ,
,  , ,
, ,  and
and
Abstract
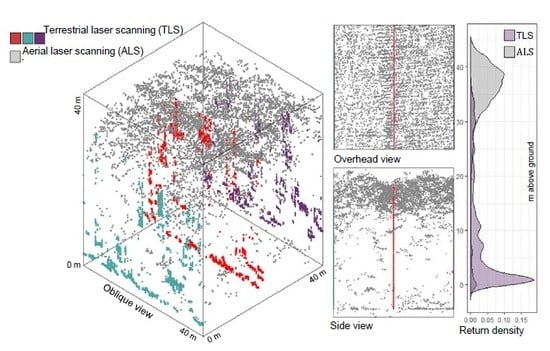
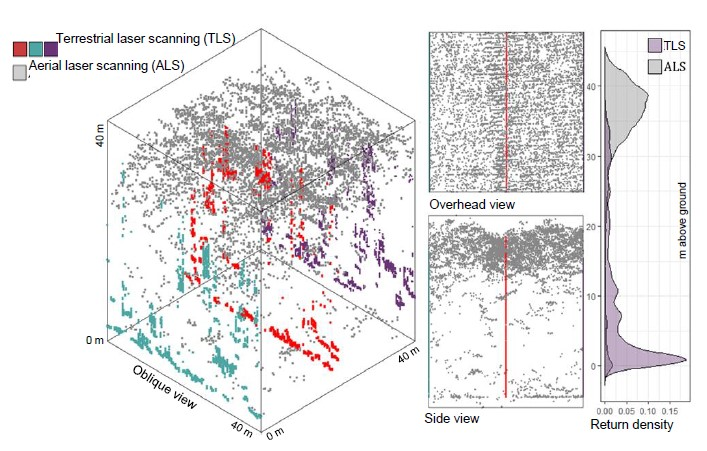
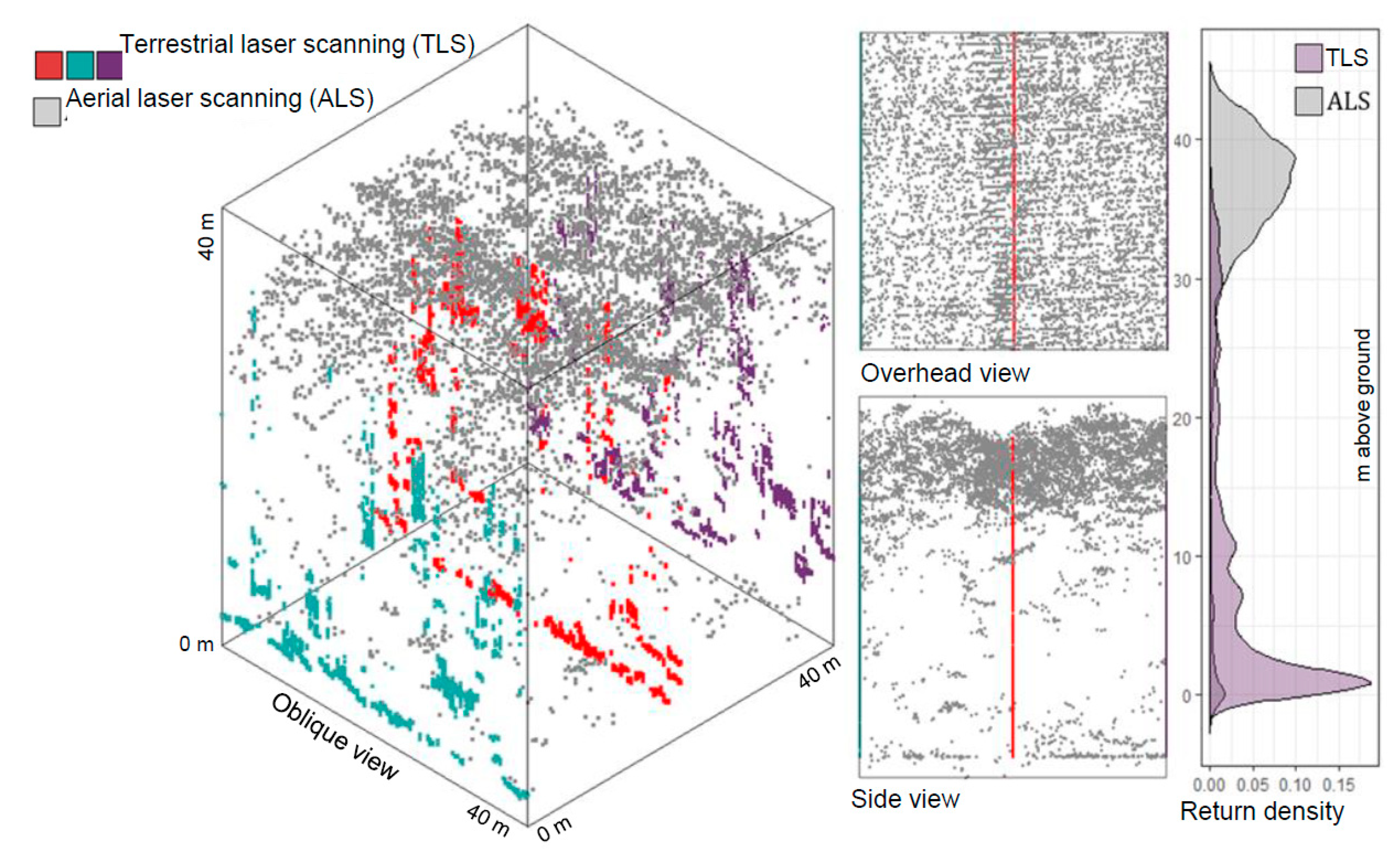
1. Introduction
2. Materials and Methods
2.1. LiDAR Data Collection and Structural Diversity Metrics
2.2. Data Analysis
3. Results
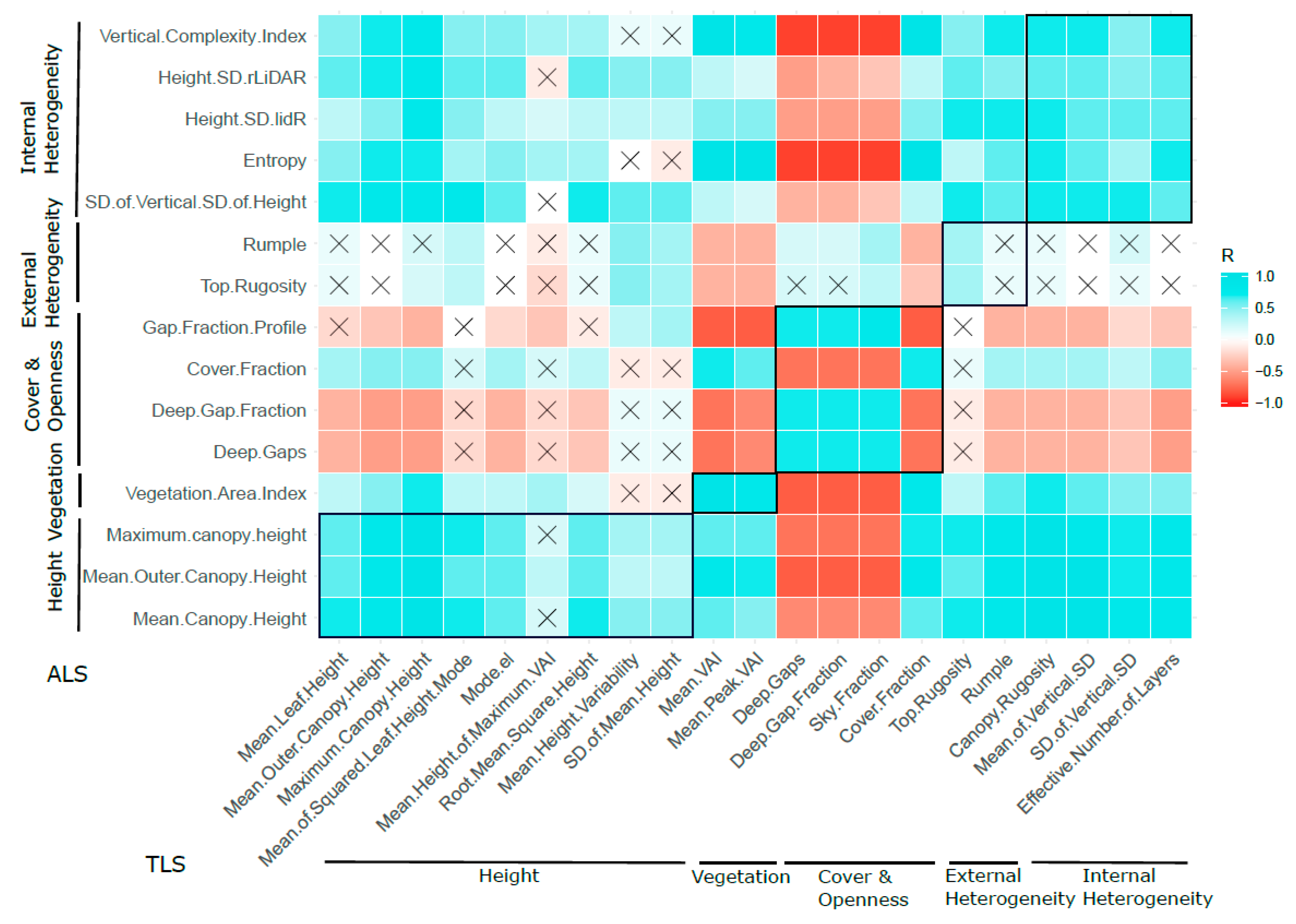
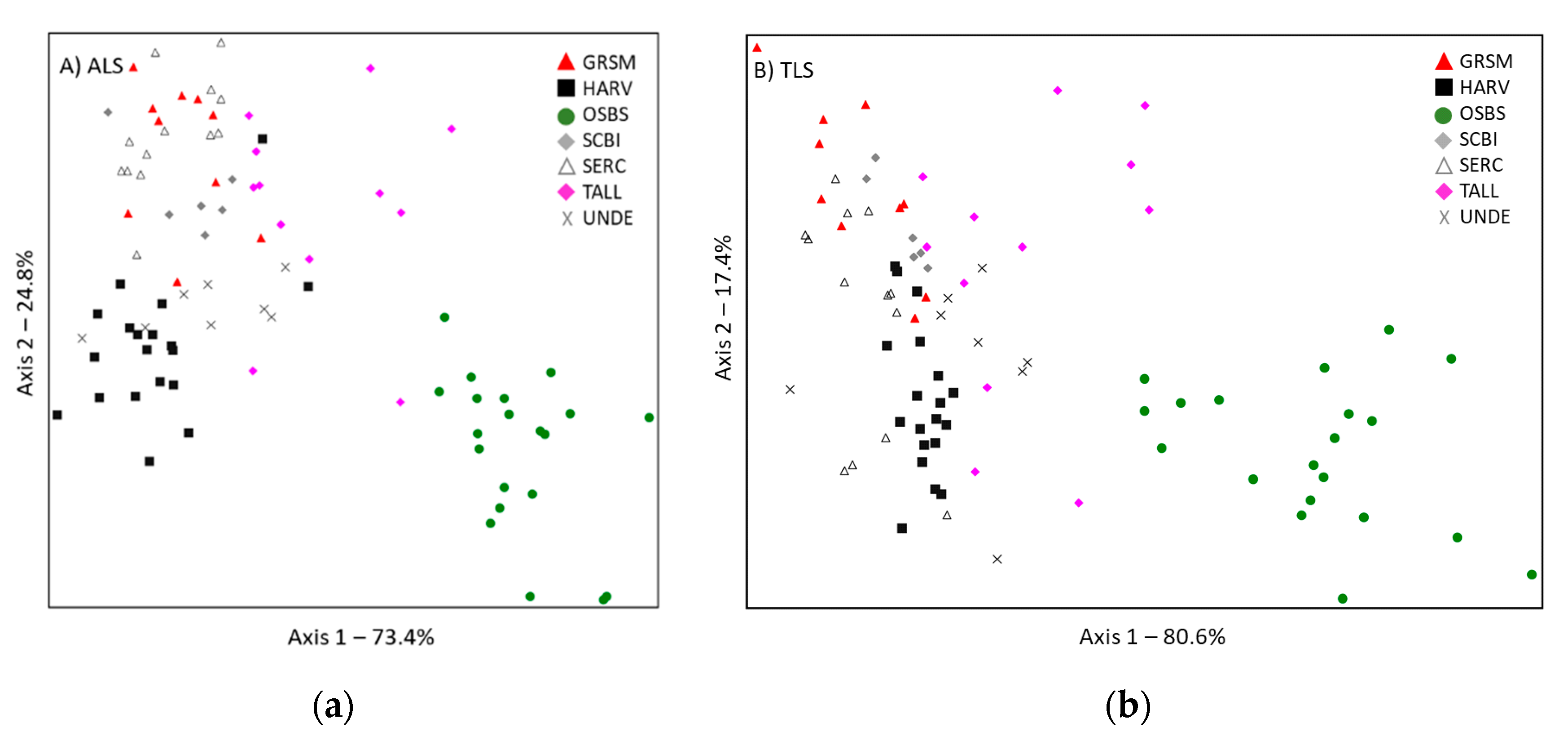
3.1. Univariate Comparison of ALS and TLS Metrics
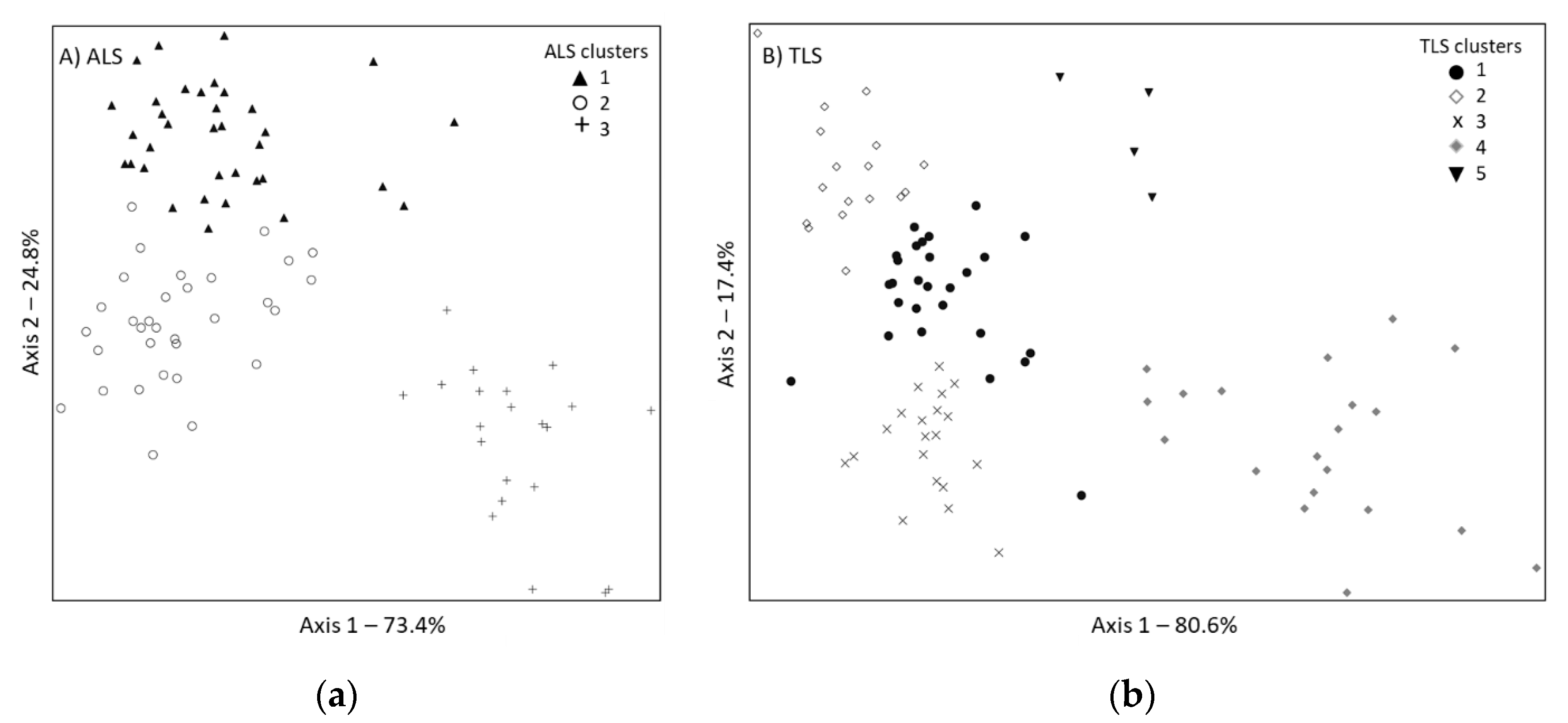
3.2. Multivariate Comparison of ALS and TLS Classifications
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- LaRue, E.A.; Hardiman, B.S.; Elliott, J.M.; Fei, S. Structural diversity as a predictor of ecosystem function. Environ. Res. Lett. 2019, 14, 114011. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bohrer, G.; Fahey, R.T.; Hardiman, B.S.; Morin, T.H.; Stovall, A.E.L.; Gough, C.M. Quantifying vegetation and canopy structural complexity from TLS data using the forestr r package. Methods Ecol. Evol. 2018, 9, 2057–2066. [Google Scholar] [CrossRef]
- Niinemets, U. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef] [PubMed]
- Matheny, A.M.; Bohrer, G.; Garrity, S.R.; Morin, T.H.; Howard, C.J.; Vogel, C.S. Observations of stem water storage in trees of opposing hydraulic strategies. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Gough, C.M.; Halperin, A.; Hofmeister, K.L.; Nave, L.E.; Bohrer, G.; Curtis, P.S. Maintaining high rates of carbon storage in old forests: A mechanism linking canopy structure to forest function. For. Ecol. Manag. 2013, 298, 111–119. [Google Scholar] [CrossRef]
- Fotis, A.T.; Morin, T.H.; Fahey, R.T.; Hardiman, B.S.; Bohrer, G.; Curtis, P.S. Forest structure in space and time: Biotic and abiotic determinants of canopy complexity and their effects on net primary productivity. Agric. For. Meteorol. 2018, 250, 181–191. [Google Scholar] [CrossRef]
- Fahey, R.T.; Atkins, J.W.; Gough, C.M.; Hardiman, B.S.; Nave, L.E.; Tallant, J.M.; Nadehoffer, K.J.; Vogel, C.; Scheuermann, C.M.; Stuart-Haëntjens, E.; et al. Defining a spectrum of integrative trait-based vegetation canopy structural types. Ecol. Lett. 2019, 22, 2049–2059. [Google Scholar] [CrossRef]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bohrer, G.; Fahey, R.T.; Hardiman, B.S.; Gough, C.M.; Morin, T.H.; Stovall, A.; Zimmerman, N. Forestr: Ecosystem and Canopy Structural Complexity Metrics from LiDAR; R Package Version 1.0.1; 2018; Available online: https://cran.r-project.org/web/packages/forestr/ (accessed on 1 January 2018).
- Wilkes, P.; Disney, M.; Vicari, M.B.; Calders, K.; Burt, A. Estimating urban above ground biomass with multi-scale LiDAR. Carbon Balanc. Manag. 2018, 13, 10. [Google Scholar] [CrossRef]
- Fisher, R.A.; Koven, C.D.; Anderegg, W.R.L.; Christoffersen, B.O.; Dietze, M.C.; Farrior, C.E.; Moorcroft, P.R. Vegetation demographics in Earth System Models: A review of progress and priorities. Glob. Chang. Biol. 2018, 24, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Shiklomanov, A.N.; Bradley, B.A.; Dahlin, K.M.; M Fox, A.; Gough, C.M.; Hoffman, F.M.; M Middleton, E.; Serbin, S.P.; Smallman, L.; Smith, W.K. Enhancing global change experiments through integration of remote-sensing techniques. Front. Ecol. Environ. 2019, 17, 215–224. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Harding, D.J.; Parker, G.G.; Acker, S.A.; Gower, S.T. LiDAR remote sensing of above-ground biomass in three biomes. Glob. Ecol. Biogeogr. 2002, 11, 393–399. [Google Scholar] [CrossRef]
- Lim, K.; Treitz, P.; Wulder, M.; St-Onge, B.; Flood, M. LiDAR remote sensing of forest structure. Prog. Phys. Geogr. Earth Environ. 2003, 1, 88–106. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Hutyra, L.R.; Hardiman, B.S.; Raciti, S.M. Characterizing forest structure variations across an intact tropical peat dome using field samplings and airborne LiDAR. Ecol. Appl. 2016, 26, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Disney, M.I.; Boni Vicari, M.; Burt, A.; Calders, K.; Lewis, L.; Raumonen, P.; Wilkes, P. Weighing trees with lasers: Advances, challenges and opportunities. Interface Focus 2018, 8, 20170048. [Google Scholar] [CrossRef]
- LaRue, E.A.; Atkins, J.W.; Dahlin, K.; Fahey, R.; Fei, S.; Gough, C.; Hardirnan, B.S. Linking Landsat to terrestrial LiDAR: Vegetation metrics of forest greenness are correlated with canopy structural complexity. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 420–427. [Google Scholar] [CrossRef]
- Paynter, I.; Genest, D.; Saenz, E.; Peri, F.; Boucher, P.; Li, Z.; Strahler, A.; Schaaf, C. Classifying ecosystems with metaproperties from terrestrial laser scanner data. Methods Ecol. Evol. 2018, 9, 210–222. [Google Scholar] [CrossRef]
- Newnham, G.J.; Armston, J.D.; Calders, K.; Disney, M.I.; Lovell, J.L.; Schaaf, C.B.; Strahler, A.H.; Danson, F.M. Terrestrial Laser Scanning for Plot-Scale Forest Measurement. Curr. For. Rep. 2015, 1, 239–251. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. LiDAR remote sensing for ecosystem studies. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Parker, G.G.; Harding, D.J.; Berger, M.L. A portable LiDAR system for rapid determination of forest canopy structure. J. Appl. Ecol. 2004, 41, 755–767. [Google Scholar] [CrossRef]
- Hilker, T.; Coops, N.C.; Newnham, G.J.; van Leeuwen, M.; Wulder, M.A.; Stewart, J.; Culvenor, D.S. Comparison of terrestrial and airborne LiDAR in describing stand structure of a thinned lodgepole pine forest. J. For. 2012, 110, 97–104. [Google Scholar] [CrossRef]
- Wing, B.M.; Ritchie, M.W.; Boston, K.; Cohen, W.B.; Gitelman, A.; Olsen, M.J. Prediction of understory vegetation cover with airborne lidar in an interior ponderosa pine forest. Remote Sens. Environ. 2012, 124, 730–741. [Google Scholar] [CrossRef]
- Ferraz, A.; Bretar, F.; Jacquemoud, S.; Gonçalves, G.; Pereira, L.; Tomé, M.; Soares, P. 3-D mapping of a multi-layered Mediterranean forest using ALS data. Remote Sens. Environ. 2012, 121, 210–223. [Google Scholar] [CrossRef]
- Campbell, M.J.; Dennison, P.E.; Hudak, A.T.; Parham, L.M.; Butler, B.W. Quantifying understory vegetation density using small-footprint airborne lidar. Remote Sens. Environ. 2018, 215, 330–342. [Google Scholar] [CrossRef]
- Venier, L.A.; Swystun, T.; Mazerolle, M.J.; Kreutzweiser, D.P.; Wainio-Keizer, K.L.; McIlwrick, K.A.; Woods, M.E.; Wang, X. Modelling vegetation understory cover using LiDAR metrics. PLoS ONE 2019, 14, e0220096. [Google Scholar] [CrossRef]
- Hamraz, H.; Contreras, M.A.; Zhang, J. Forest understory trees can be segmented accurately within sufficiently dense airborne laser scanning point clouds. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Hamraz, H.; Contreras, M.A.; Zhang, J. Vertical stratification of forest canopy for segmentation of understory trees within small-footprint airborne LiDAR point clouds. ISPRS J. Photogramm. Remote Sens. 2017, 130, 385–392. [Google Scholar] [CrossRef]
- Kankare, V.; Vauhkonen, J.; Tanhuanpää, T.; Holopainen, M.; Vastaranta, M.; Joensuu, M.; Krooks, A.; Hyyppä, J.; Hyyppä, H.; Alho, P.; et al. Accuracy in estimation of timber assortments and stem distribution—A comparison of airborne and terrestrial laser scanning techniques. ISPRS J. Photogramm. Remote Sens. 2014, 97, 89–97. [Google Scholar] [CrossRef]
- Trochta, J.; Kruček, M.; Vrška, T.; Kraâl, K. 3D Forest: An application for descriptions of three-dimensional forest structures using terrestrial LiDAR. PLoS ONE 2017, 12, e0176871. [Google Scholar] [CrossRef]
- Wang, Y.; Lehtomäki, M.; Liang, X.; Pyörälä, J.; Kukko, A.; Jaakkola, A.; Liu, J.; Feng, Z.; Chen, R.; Hyyppä, J. Is field-measured tree height as reliable as believed—A comparison study of tree height estimates from field measurement, airborne laser scanning and terrestrial laser scanning in a boreal forest. ISPRS J. Photogramm. Remote Sens. 2019, 147, 132–145. [Google Scholar] [CrossRef]
- Liang, X.; Wang, Y.; Pyörälä, J.; Lehtomäki, M.; Yu, X.; Kaartinen, H.; Kukko, A.; Honkavaara, E.; Issaoui, A.E.I.; Nevalainen, O.; et al. Forest in situ observations using unmanned aerial vehicle as an alternative of terrestrial measurements. For. Ecosyst. 2019, 6, 20. [Google Scholar] [CrossRef]
- Listopad, C.M.C.S.; Drake, J.B.; Masters, R.E.; Weishampel, J.F. Portable and airborne small footprint LiDAR: Forest canopy structure estimation of fire managed plots. Remote Sens. 2011, 3, 1284–1307. [Google Scholar] [CrossRef]
- Hopkinson, C.; Lovell, J.; Chasmer, L.; Jupp, D.; Kljun, N.; van Gorsel, E. Integrating terrestrial and airborne LiDAR to calibrate a 3D canopy model of effective leaf area index. Remote Sens. Environ. 2013, 136, 301–314. [Google Scholar] [CrossRef]
- Hilker, T.; van Leeuwen, M.; Coops, N.C.; Wulder, M.A.; Newnham, G.J.; Jupp, D.L.B.; Culvenor, D.S. Comparing canopy metrics derived from terrestrial and airborne laser scanning in a Douglas-fir dominated forest stand. Trees Struct. Funct. 2010, 24, 819–832. [Google Scholar] [CrossRef]
- Braun, L. Deciduous Forests of Eastern North America; Hafner: New York, NY, USA, 1950. [Google Scholar]
- Atkins, J.W.; Fahey, R.T.; Hardiman, B.S.; Gough, C.M. Forest Canopy Structural Complexity and Light Absorption Relationships at the Subcontinental Scale. J. Geophys. Res. Biogeosci. 2018, 123, 1387–1405. [Google Scholar] [CrossRef]
- Team, R.D.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 30 July 2018).
- National Ecological Observatory Network. Data Products: DP1.30003.001; Battelle: Boulder, CO, USA, 2019; Available online: http://data.neonscience.org (accessed on 1 January 2019).
- Roussel, J.R.; Auty, D. LidR: Airborne LiDAR Data Manipulation and Visualization for Forestry Applications; R Package Version, Version 1.3.1; 2018; Available online: https://cran.r-project.org/web/packages/lidR (accessed on 30 July 2018).
- Silva, C.A.; Crookston, N.L.; Hudak, A.T.; Vierling, L.A.; Klauberg, C.; Cardil, A. rLiDAR: LiDAR Data Processing and Visualization; R Package Version 0.1.1; 2017; Available online: https://cran.r-project.org/web/packages/rLiDAR (accessed on 30 August 2018).
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 5.0 for Windows; MjM Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Mccune, B.; Grace, J. Analysis of Ecological Communities; MJM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Meadows, M.E.; Hill, T.R. Issues in Biogeography: Diversity in theory and practice. S. Afr. Geogr. J. 2012, 6245, 116–124. [Google Scholar] [CrossRef]
- Paynter, I.; Saenz, E.; Genest, D.; Peri, F.; Erb, A.; Li, Z.; Wiggin, K.; Muir, J.; Raumonen, P.; Schaaf, E.S.; et al. Observing ecosystems with lightweight, rapid-scanning terrestrial lidar scanners. Remote Sens. Ecol. Conserv. 2016, 2, 174–189. [Google Scholar] [CrossRef]
- Mund, J.-P.; Wilke, R.; Körner, M.; Schultz, A. Detecting Multi-layered Forest Stands Using High Density Airborne LiDAR Data. J. Geogr. Inf. Sci. 2015, 1, 178–188. [Google Scholar] [CrossRef]
- Donager, J.J.; Sankey, T.T.; Sankey, J.B.; Sanchez Meador, A.J.; Springer, A.E.; Bailey, J.D. Examining forest structure with terrestrial lidar: Suggestions and novel techniques based on comparisons between scanners and forest treatments. Earth Space Sci. 2018, 5, 753–776. [Google Scholar] [CrossRef]
- Gough, C.M.; Atkins, J.W.; Fahey, R.T.; Hardiman, B.S. High rates of primary production in structurally complex forests. Ecology 2019, 100, e02864. [Google Scholar] [CrossRef] [PubMed]
- Kleindl, W.; Stoy, P.; Binford, M.W.; Desai, A.R.; Dietze, M.C.; Schultz, C.A.; Starr, G.; Staudhammer, C.L.; Wood David, J.A. Toward a Social-Ecological Theory of Forest Macrosystems for Improved Ecosystem Management. Forests 2018, 9, 200. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (ID) | Ecoclimatic Domain | Dominant Forest Type | NPlots |
|---|---|---|---|
| Harvard Forest (HARV) | Northeast | Mixed temperate | 19 |
| Smithsonian Conservation Biology Institute (SCBI) | Mid-Atlantic | Mixed temperate | 6 |
| Smithsonian Environmental Research Center (SERC) | Mid-Atlantic | Temperate deciduous | 13 |
| Ordway-Swisher Biological Station (OSBS) | Southeast | Pine Savannah | 20 |
| University of Notre Dame Environmental Research Center East (UNDE) | Great Lakes | Mixed temperate | 8 |
| Great Smoky Mountain National Park (GRSM) | Appalachians and Cumberland Plateau | Temperate rainforest | 10 |
| Talladega National Forest (TALL) | Ozarks Complex | Pine savannah | 12 |
| Category | ALS Metric | TLS Metric | |
|---|---|---|---|
| Height | Mean canopy height (H), Mean outer canopy height (MOCH), Maximum canopy height (Hmax) | Mean leaf height (Mean H), Mean outer canopy height (MOCH), Maximum canopy height (Max.can.ht), Mean of squared leaf height model (Mode.2), Mode.el (Model.el), Mean height of maximum VAI (Max.el), Root mean square height (meanHRMS), Mean height variability (meanHvar), SD of mean height (Height.2) | |
| Cover and openness | Deep gaps (DG), Deep gap fraction (DGF), Cover fraction (CF), Gap fraction profile (GFP) | Deep gaps (DG), Deep gap fraction (DGF), Sky fraction (SF), Cover fraction (CF) | |
| Heterogeneity | External | Top rugosity (TR), Rumple (Rumple) | Top rugosity (TR), Rumple (Rumple) |
| Internal | SD of vertical SD of height (StdStd), Entropy (Entropy), Height SD (HSD: rLiDAR), Height SD (StdH: lidR), Vertical complexity index (VCI) | Canopy rugosity (Rugosity), Mean of vertical SD (MeanStd), SD of vertical SD (StdStd), Effective number of layers (ENL) | |
| Vegetation area | Vegetation area index (VAI) | Mean VAI (Mean.VAI), Mean peak VAI (Mean.peak.VAI) | |
| System Specifications | Optech ALTM Gemini (ALS) | Riegl LD90-3100VHS-FLP (TLS) |
|---|---|---|
| Returns per Pulse | Four | Five |
| Wavelength | 1064 nm | 900 nm |
| Measurement Range | 150–4000 m | 60 m (ρ ≥ 0.1)–200 m (ρ ≥ 0.8) |
| Range Accuracy (typical) | ± 5–30 cm | ±2.5 cm |
| Bean Divergence Angle | 0.25 mrad × 0.8 mrad | 3 mrad × 5 mrad |
| Measurement Rate (per second) | 0–70 (programmable) | 2000 |
| Average Point Density | 1–4 points per m2 | 500–2000 points per linear meter |
| Laser Product Classification | Class IV (US FDA 21 CFR) | IEC 60825–1:2007 (Eye-safe) |
| Category | ALS | TLS | r |
|---|---|---|---|
| Height | MOCH | MOCH | 0.80 * |
| H | H | 0.72 * | |
| Hmax | Max.can.ht | 0.92 * | |
| Cover and openness | DGF | DGF | 0.66 * |
| CF | CF | 0.74 * | |
| External heterogeneity | Rumple | Rumple | 0.09 |
| TR | TR | 0.44 * | |
| Internal heterogeneity | StdH | MeanStd | 0.61 * |
| StdStd | Rugosity | 0.74 * | |
| Vegetation area | VAI | VAI | 0.87 * |
| Cluster | TLS1 | TLS2 | TLS3 | TLS4 | TLS5 | Total |
|---|---|---|---|---|---|---|
| ALS1 | 12 | 15 | 4 | 0 | 4 | 35 |
| ALS2 | 14 | 2 | 16 | 0 | 0 | 32 |
| ALS3 | 1 | 0 | 0 | 20 | 0 | 21 |
| Total | 27 | 17 | 20 | 20 | 4 | 88 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaRue, E.A.; Wagner, F.W.; Fei, S.; Atkins, J.W.; Fahey, R.T.; Gough, C.M.; Hardiman, B.S. Compatibility of Aerial and Terrestrial LiDAR for Quantifying Forest Structural Diversity. Remote Sens. 2020, 12, 1407. https://doi.org/10.3390/rs12091407
LaRue EA, Wagner FW, Fei S, Atkins JW, Fahey RT, Gough CM, Hardiman BS. Compatibility of Aerial and Terrestrial LiDAR for Quantifying Forest Structural Diversity. Remote Sensing. 2020; 12(9):1407. https://doi.org/10.3390/rs12091407
Chicago/Turabian StyleLaRue, Elizabeth A., Franklin W. Wagner, Songlin Fei, Jeff W. Atkins, Robert T. Fahey, Christopher M. Gough, and Brady S. Hardiman. 2020. "Compatibility of Aerial and Terrestrial LiDAR for Quantifying Forest Structural Diversity" Remote Sensing 12, no. 9: 1407. https://doi.org/10.3390/rs12091407
APA StyleLaRue, E. A., Wagner, F. W., Fei, S., Atkins, J. W., Fahey, R. T., Gough, C. M., & Hardiman, B. S. (2020). Compatibility of Aerial and Terrestrial LiDAR for Quantifying Forest Structural Diversity. Remote Sensing, 12(9), 1407. https://doi.org/10.3390/rs12091407