
Mineralocorticoid Receptor Signaling in Peripheral Blood Cells in Patients with Multiple Sclerosis

Abstract
:1. Introduction
2. Results
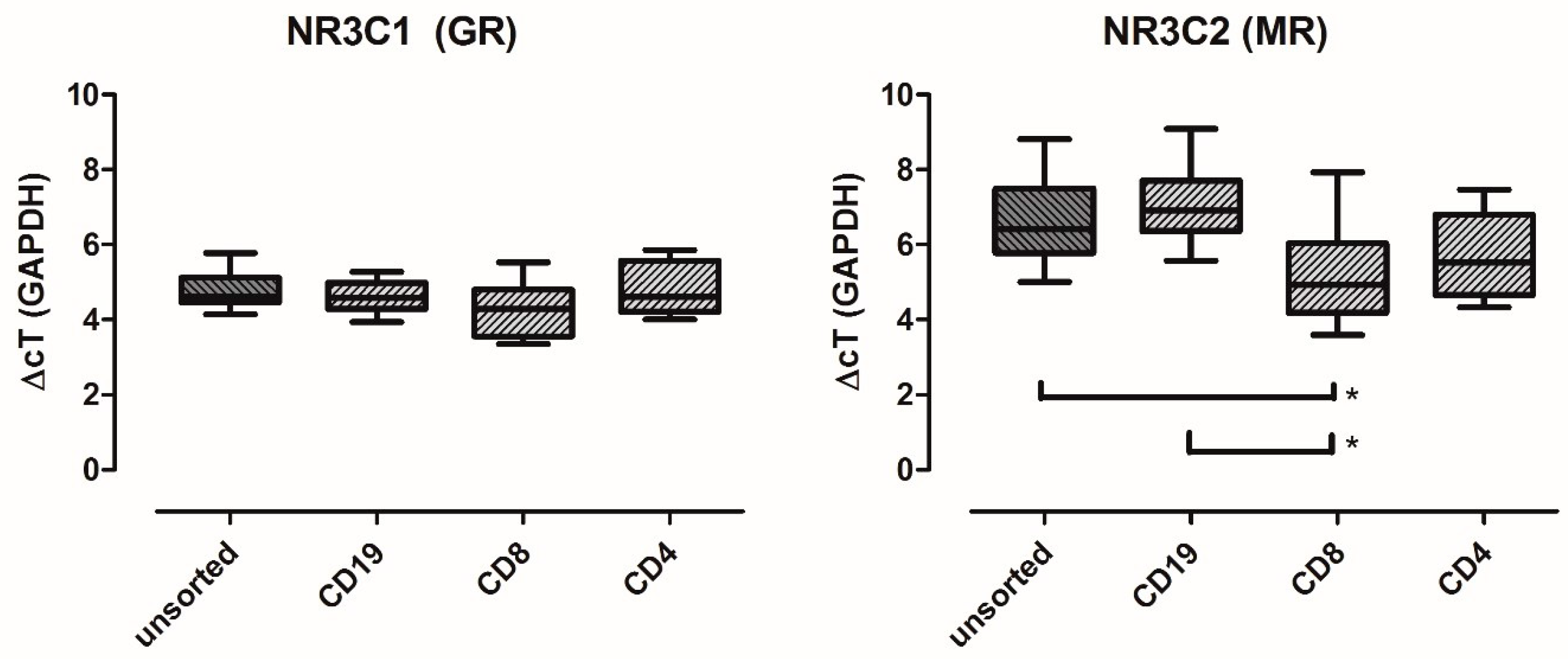
2.1. MR Is Expressed in Major PBMC Subsets
2.2. Ovarian Tumor Domain-Containing DUB 1 (OTUD1) Is a Marker for MR Signaling in PBMCs
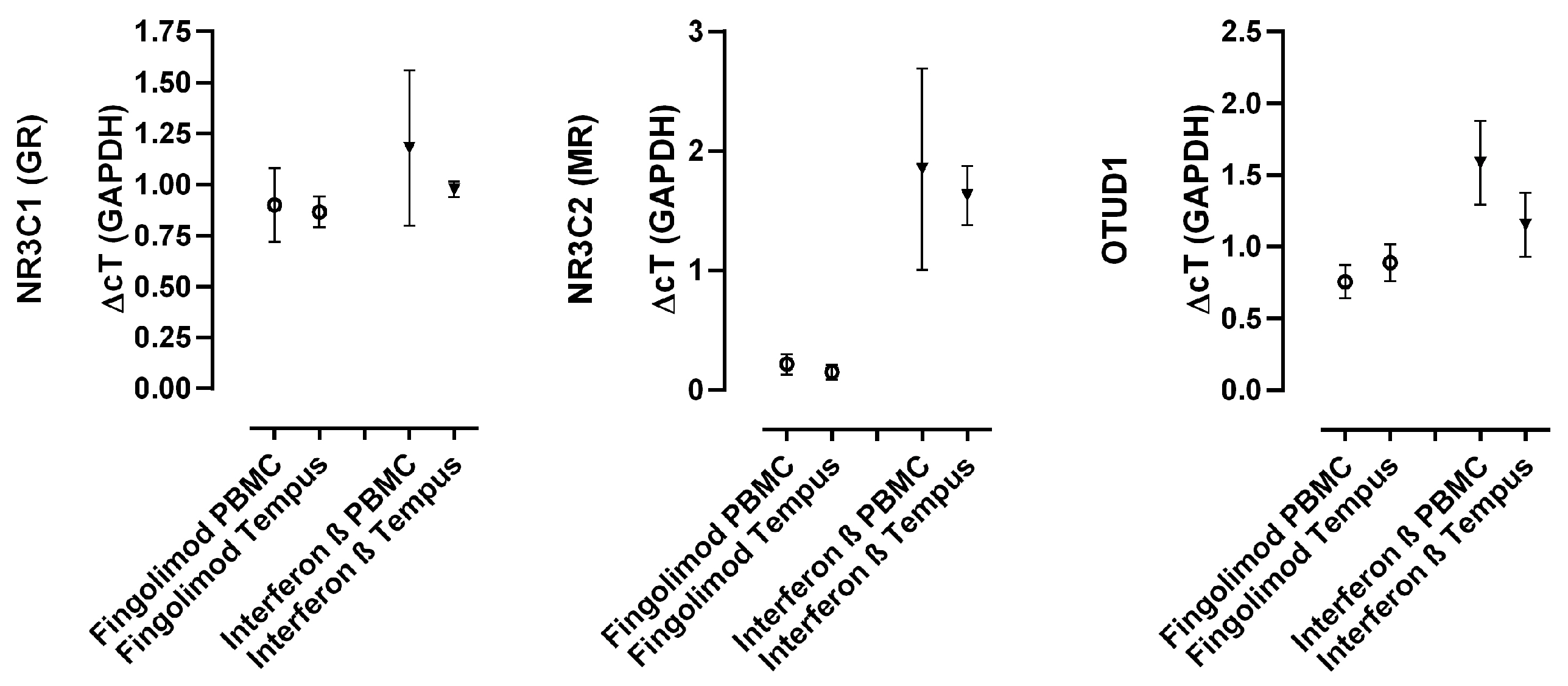
2.3. MR and OTUD1 Expression Can Reliably Be Measured in Lysed Whole Blood Samples
2.4. In Multiple Sclerosis, the Expression of Both MR and OTUD1 Is Altered
2.5. Disease-Modifying Treatments Differentially Affect MR and OTUD1 Expression
3. Discussion
3.1. Relevance of Detected MR Signaling and OTUD1 in the Context of Autoimmunity
3.2. The Search for MR Marker Genes Relevant In Vivo
3.3. Role of MR Signaling in MS
3.4. Future Aims and Perspectives
4. Conclusions
5. Materials and Methods
5.1. Subjects
5.2. Clinical Assessment of Patients’ Disability (EDSS)
5.3. Cell Culture
5.4. RNA Isolation and cDNA Synthesis
5.5. Gene Expression Analysis (Microarray)
5.6. Quantitative Real-Time PCR
5.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramagopalan, S.V.; Sadovnick, A.D. Epidemiology of Multiple Sclerosis. Neurol. Clin. 2011, 29, 207–217. [Google Scholar] [CrossRef]
- International Multiple Sclerosis Genetics Consortium; Wellcome Trust Case Control Consortium 2; Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.A.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Epidemiology in multiple sclerosis: A pilgrim’s progress. Brain 2013, 136, 2904–2917. [Google Scholar] [CrossRef] [PubMed]
- Legroux, L.; Arbour, N. Multiple Sclerosis and T Lymphocytes: An Entangled Story. J. Neuroimmune Pharmacol. 2015, 10, 528–546. [Google Scholar] [CrossRef] [PubMed]
- McFarland, H.F.; Martin, R. Multiple sclerosis: A complicated picture of autoimmunity. Nat. Immunol. 2007, 8, 913–919. [Google Scholar] [CrossRef]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Rohleder, N.; Eisenhofer, G.; Lange, J.; Ziemssen, T. Time matters—Acute stress response and glucocorticoid sensitivity in early multiple sclerosis. Brain Behav. Immun. 2014, 41, 82–89. [Google Scholar] [CrossRef]
- Kümpfel, T.; Schwan, M.; Weber, F.; Holsboer, F.; Trenkwalder, C.; Then Bergh, F. Hypothalamo-pituitary-adrenal axis activity evolves differentially in untreated versus treated multiple sclerosis. Psychoneuroendocrinology 2014, 45, 87–95. [Google Scholar] [CrossRef]
- Schumann, E.M.; Kumpfel, T.; Then Bergh, F.; Trenkwalder, C.; Holsboer, F.; Auer, D.P. Activity of the hypothalamic-pituitary-adrenal axis in multiple sclerosis: Correlations with gadolinium-enhancing lesions and ventricular volume. Ann. Neurol. 2002, 51, 763–767. [Google Scholar] [CrossRef] [PubMed]
- DeRijk, R.H.; Eskandari, F.; Sternberg, E.M. Corticosteroid resistance in a subpopulation of multiple sclerosis patients as measured by ex vivo dexamethasone inhibition of LPS induced IL-6 production. J. Neuroimmunol. 2004, 151, 180–188. [Google Scholar] [CrossRef] [PubMed]
- van Winsen, L.M.L.; Muris, D.F.R.; Polman, C.H.; Dijkstra, C.D.; van den Berg, T.K.; Uitdehaag, B.M.J. Sensitivity to glucocorticoids is decreased in relapsing remitting multiple sclerosis. J. Clin. Endocrinol. Metab. 2005, 90, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Ysrraelit, M.C.; Gaitan, M.I.; Lopez, A.S.; Correale, J. Impaired hypothalamic-pituitary-adrenal axis activity in patients with multiple sclerosis. Neurology 2008, 71, 1948–1954. [Google Scholar] [CrossRef]
- Evangelopoulos, M.E.; Nasiri-Ansari, N.; Kassi, E.; Papadopoulou, A.; Evangelopoulos, D.S.; Moutsatsou, P. Methylprednisolone stimulated gene expression (GILZ, MCL-1) and basal cortisol levels in multiple sclerosis patients in relapse are associated with clinical response. Sci. Rep. 2021, 11, 19462. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.; Busse, K.; Stoppe, M.; Cotte, S.; Ettrich, B.; Then Bergh, F. Corticosteroid receptor expression and in vivo glucocorticoid sensitivity in multiple sclerosis. J. Neuroimmunol. 2014, 276, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Hasan, S.; Sharma, S.; Nagra, S.; Yamaguchi, D.T.; Wong, D.T.W.; Hahn, B.H.; Hossain, A. Th17 cells in inflammation and autoimmunity. Autoimmun. Rev. 2014, 13, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Kleinewietfeld, M.; Manzel, A.; Titze, J.; Kvakan, H.; Yosef, N.; Linker, R.A.; Muller, D.N.; Hafler, D.A. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature 2013, 496, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Hucke, S.; Eschborn, M.; Liebmann, M.; Herold, M.; Freise, N.; Engbers, A.; Ehling, P.; Meuth, S.G.; Roth, J.; Kuhlmann, T.; et al. Sodium chloride promotes pro-inflammatory macrophage polarization thereby aggravating CNS autoimmunity. J. Autoimmun. 2016, 67, 90–101. [Google Scholar] [CrossRef]
- Platten, M.; Youssef, S.; Hur, E.M.; Ho, P.P.; Han, M.H.; Lanz, T.V.; Phillips, L.K.; Goldstein, M.J.; Bhat, R.; Raine, C.S.; et al. Blocking angiotensin-converting enzyme induces potent regulatory T cells and modulates TH1- and TH17-mediated autoimmunity. Proc. Natl. Acad. Sci. USA 2009, 106, 14948–14953. [Google Scholar] [CrossRef]
- Farez, M.F.; Fiol, M.P.; Gaitan, M.I.; Quintana, F.J.; Correale, J. Sodium intake is associated with increased disease activity in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 26–31. [Google Scholar] [CrossRef]
- Fitzgerald, K.C.; Munger, K.L.; Hartung, H.-P.; Freedman, M.S.; Montalbán, X.; Edan, G.; Wicklein, E.-M.; Radue, E.-W.; Kappos, L.; Pohl, C.; et al. Sodium intake and multiple sclerosis activity and progression in BENEFIT. Ann. Neurol. 2017, 82, 20–29. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.; Graves, J.; Waldman, A.; Lotze, T.; Schreiner, T.; Belman, A.; Greenberg, B.; Weinstock-Guttman, B.; Aaen, G.; Tillema, J.-M.; et al. A case-control study of dietary salt intake in pediatric-onset multiple sclerosis. Mult. Scler. Relat. Disord. 2016, 6, 87–92. [Google Scholar] [CrossRef]
- Nourbakhsh, B.; Graves, J.; Casper, T.C.; Lulu, S.; Waldman, A.; Belman, A.; Greenberg, B.; Weinstock-Guttman, B.; Aaen, G.; Tillema, J.-M.; et al. Dietary salt intake and time to relapse in paediatric multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1350–1353. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Phung, Q.; Chan, S.; Chaudhari, R.; Quan, C.; O’Rourke, K.M.; Eby, M.; Pietras, E.; Cheng, G.; Bazan, J.F.; et al. DUBA: A deubiquitinase that regulates type I interferon production. Science 2007, 318, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, D.; Wang, P.; Zhao, Y.; You, F. OTUD1 Negatively Regulates Type I IFN Induction by Disrupting Noncanonical Ubiquitination of IRF3. J. Immunol. 2020, 204, 1904–1918. [Google Scholar] [CrossRef]
- Lu, D.; Song, J.; Sun, Y.; Qi, F.; Liu, L.; Jin, Y.; McNutt, M.A.; Yin, Y. Mutations of deubiquitinase OTUD1 are associated with autoimmune disorders. J. Autoimmun. 2018, 94, 156–165. [Google Scholar] [CrossRef]
- Wu, B.; Qiang, L.; Zhang, Y.; Fu, Y.; Zhao, M.; Lei, Z.; Lu, Z.; Wei, Y.-G.; Dai, H.; Ge, Y.; et al. The deubiquitinase OTUD1 inhibits colonic inflammation by suppressing RIPK1-mediated NF-κB signaling. Cell. Mol. Immunol. 2022, 19, 276–289. [Google Scholar] [CrossRef]
- Cheng, Q.; Wang, D.; Lai, X.; Liu, Y.; Zuo, Y.; Zhang, W.; Lei, L.; Chen, J.; Liu, H.; Wang, Y.; et al. The OTUD1-Notch2-ICD axis orchestrates allogeneic T cell-mediated graft-versus-host disease. Blood 2023, 141, 1474–1488. [Google Scholar] [CrossRef]
- Liu, Q.; Mallette, E.; Zheng, H.; Zhang, W. Development of an OTUD1 ubiquitin variant inhibitor. Biochem. J. 2023, 480, 1317–1330. [Google Scholar] [CrossRef]
- Yang, J.; Fuller, P.J. Interactions of the mineralocorticoid receptor—Within and without. Mol. Cell Endocrinol. 2012, 350, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Le Tallec, L.; Lombès, M. The mineralocorticoid receptor: A journey exploring its diversity and specificity of action. Mol. Endocrinol. 2005, 19, 2211–2221. [Google Scholar] [CrossRef] [PubMed]
- Fuller, P.J.; Lim-Tio, S.S.; Brennan, F.E. Specificity in mineralocorticoid versus glucocorticoid action. Kidney Int. 2000, 57, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Bene, N.C.; Alcaide, P.; Wortis, H.H.; Jaffe, I.Z. Mineralocorticoid receptors in immune cells: Emerging role in cardiovascular disease. Steroids 2014, 91, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, J.; Bajaj, T.; Klengel, C.; Chatzinakos, C.; Ebert, T.; Dedic, N.; McCullough, K.M.; Lardenoije, R.; Joëls, M.; Meijer, O.C.; et al. Mineralocorticoid receptors dampen glucocorticoid receptor sensitivity to stress via regulation of FKBP5. Cell Rep. 2021, 35, 109185. [Google Scholar] [CrossRef]
- Shibata, S. 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: Mineralocorticoid receptor and NaCl transport mechanisms in the renal distal nephron. J. Endocrinol. 2017, 234, T35–T47. [Google Scholar] [CrossRef]
- Alvarez Quintero, G.S.; Lima, A.; Roig, P.; Meyer, M.; de Kloet, E.R.; de Nicola, A.F.; Garay, L.I. Effects of the mineralocorticoid receptor antagonist eplerenone in experimental autoimmune encephalomyelitis. J. Steroid Biochem. Mol. Biol. 2024, 238, 106461. [Google Scholar] [CrossRef]
- Goel, N.; Workman, J.L.; Lee, T.T.; Innala, L.; Viau, V. Sex differences in the HPA axis. Compr. Physiol. 2014, 4, 1121–1155. [Google Scholar] [CrossRef]
- Gupta, D.; Morley, J.E. Hypothalamic-pituitary-adrenal (HPA) axis and aging. Compr. Physiol. 2014, 4, 1495–1510. [Google Scholar] [CrossRef]
- Kieseier, B.C. The Mechanism of Action of Interferon-β in Relapsing Multiple Sclerosis. CNS Drugs 2011, 25, 491–502. [Google Scholar] [CrossRef]
- Nunez, B.S.; Geng, C.-D.; Pedersen, K.B.; Millro-Macklin, C.D.; Vedeckis, W.V. Interaction between the interferon signaling pathway and the human glucocorticoid receptor gene 1A promoter. Endocrinology 2005, 146, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, B.; Kerschensteiner, M.; Korn, T. Role of the innate and adaptive immune responses in the course of multiple sclerosis. Lancet Neurol. 2015, 14, 406–419. [Google Scholar] [CrossRef]
- Biernacki, T.; Kokas, Z.; Sandi, D.; Füvesi, J.; Fricska-Nagy, Z.; Faragó, P.; Kincses, T.Z.; Klivényi, P.; Bencsik, K.; Vécsei, L. Emerging Biomarkers of Multiple Sclerosis in the Blood and the CSF: A Focus on Neurofilaments and Therapeutic Considerations. Int. J. Mol. Sci. 2022, 23, 3383. [Google Scholar] [CrossRef] [PubMed]
- Barro, C.; Healy, B.C.; Liu, Y.; Saxena, S.; Paul, A.; Polgar-Turcsanyi, M.; Guttmann, C.R.G.; Bakshi, R.; Kropshofer, H.; Weiner, H.L.; et al. Serum GFAP and NfL Levels Differentiate Subsequent Progression and Disease Activity in Patients with Progressive Multiple Sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2023, 10, e200052. [Google Scholar] [CrossRef]
- Åkesson, J.; Hojjati, S.; Hellberg, S.; Raffetseder, J.; Khademi, M.; Rynkowski, R.; Kockum, I.; Altafini, C.; Lubovac-Pilav, Z.; Mellergård, J.; et al. Proteomics reveal biomarkers for diagnosis, disease activity and long-term disability outcomes in multiple sclerosis. Nat. Commun. 2023, 14, 6903. [Google Scholar] [CrossRef] [PubMed]
- Ayrignac, X.; Le Bars, E.; Duflos, C.; Hirtz, C.; Maleska Maceski, A.; Carra-Dallière, C.; Charif, M.; Pinna, F.; Prin, P.; Menjot de Champfleur, N.; et al. Serum GFAP in multiple sclerosis: Correlation with disease type and MRI markers of disease severity. Sci. Rep. 2020, 10, 10923. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.J.; Zeidler, R.; Baber, R.; Vogel, M.; Wirkner, K.; Loeffler, M.; Ceglarek, U.; Kiess, W.; Körner, A.; Thiery, J.; et al. Reference intervals of nine steroid hormones over the life-span analyzed by LC-MS/MS: Effect of age, gender, puberty, and oral contraceptives. J. Steroid Biochem. Mol. Biol. 2019, 193, 105409. [Google Scholar] [CrossRef]
- Funder, J.W. Reconsidering the Roles of the Mineralocorticoid Receptor. Hypertension 2009, 53, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Marcus, R.; Eric, P.; Gabriel, K.R. On closed testing procedures with special reference to ordered analysis of variance. Biometrika 1976, 63, 655–660. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Controls | MS Patients | EDSS Groups | ||||||
|---|---|---|---|---|---|---|---|---|
| 0–1.5 | 2–3 | 3.5–4.5 | >5 | |||||
| n (%) | 42 | 119 | 37 (31.1) | 47 (39.5) | 24 (20.2) | 11 (9.2) | ||
| Age [years]: mean (SD)] | 33.6 (10.2) | 38.9 (10.4) | 35.0 (9.9) | 36.7 (8.9) | 45.8 (10.6) | 46.0 (7.2) | ||
| Female: n (%) | 25 (59.5) | 86 (72.3) | 28 (75.7) | 37 (78.7) | 16 (66.7) | 5 (45.5) | ||
| Disease duration [years]: mean (SD) | 6.4 (7.0) | 2.9 (3.3) | 4.8 (5.1) | 12.9 (9.1) | 11.2 (7.5) | |||
| Disease course [n] (CIS/RRMS/SPMS) | (14/99/6) | (8/29/0) | (6/41/0) | (0/23/1) | (0/6/5) | |||
| Treatment: n (%) | ||||||||
| no treatment | 42 (35.3) | 19 (51.4) | 17 (36.2) | 2 (8.3) | 4 (36.4) | |||
| interferon β | 44 (37.0) | 11 (29.7) | 14 (29.8) | 15 (62.5) | 4 (36.4) | |||
| fingolimod | 33 (27.7) | 7 (18.9) | 16 (34.0) | 7 (29.2) | 3 (27.3) | |||
| Healthy controls | MS patients | Treatment | ||||||
| no treatment | interferon β | fingolimod | ||||||
| n (%) | 42 | 119 | 42 (35.3) | 44 (37.0) | 33 (27.7) | |||
| Age [years]: mean (SD) | 33.6 (10.2) | 38.9 (10.4) | 37.6 (10.4) | 38.5 (10.8) | 41.1 (9.3) | |||
| Female: n (%) | 25 (59.5) | 86 (72.3) | 28 (66.7) | 31 (70.5) | 24 (72.7) | |||
| Disease duration [years]: mean (SD) | 6.4 (7.0) | 3.6 (5.4) | 7.5 (7.4) | 8.6 (7.3) | ||||
| Disease course [n]: (CIS/RRMS/SPMS) | (14/99/6) | (13/27/3) | (1/42/1) | (0/30/2) | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Küstermann, F.; Busse, K.; Orthgieß, J.; Stoppe, M.; Haars, S.; Then Bergh, F. Mineralocorticoid Receptor Signaling in Peripheral Blood Cells in Patients with Multiple Sclerosis. Int. J. Mol. Sci. 2024, 25, 8883. https://doi.org/10.3390/ijms25168883
Küstermann F, Busse K, Orthgieß J, Stoppe M, Haars S, Then Bergh F. Mineralocorticoid Receptor Signaling in Peripheral Blood Cells in Patients with Multiple Sclerosis. International Journal of Molecular Sciences. 2024; 25(16):8883. https://doi.org/10.3390/ijms25168883
Chicago/Turabian StyleKüstermann, Franziska, Kathy Busse, Johannes Orthgieß, Muriel Stoppe, Sarah Haars, and Florian Then Bergh. 2024. "Mineralocorticoid Receptor Signaling in Peripheral Blood Cells in Patients with Multiple Sclerosis" International Journal of Molecular Sciences 25, no. 16: 8883. https://doi.org/10.3390/ijms25168883