
Identification of Putative Candidate Genes from Galphimia spp. Encoding Enzymes of the Galphimines Triterpenoids Synthesis Pathway with Anxiolytic and Sedative Effects

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Plant Material
2.2. Transcriptome Sequencing
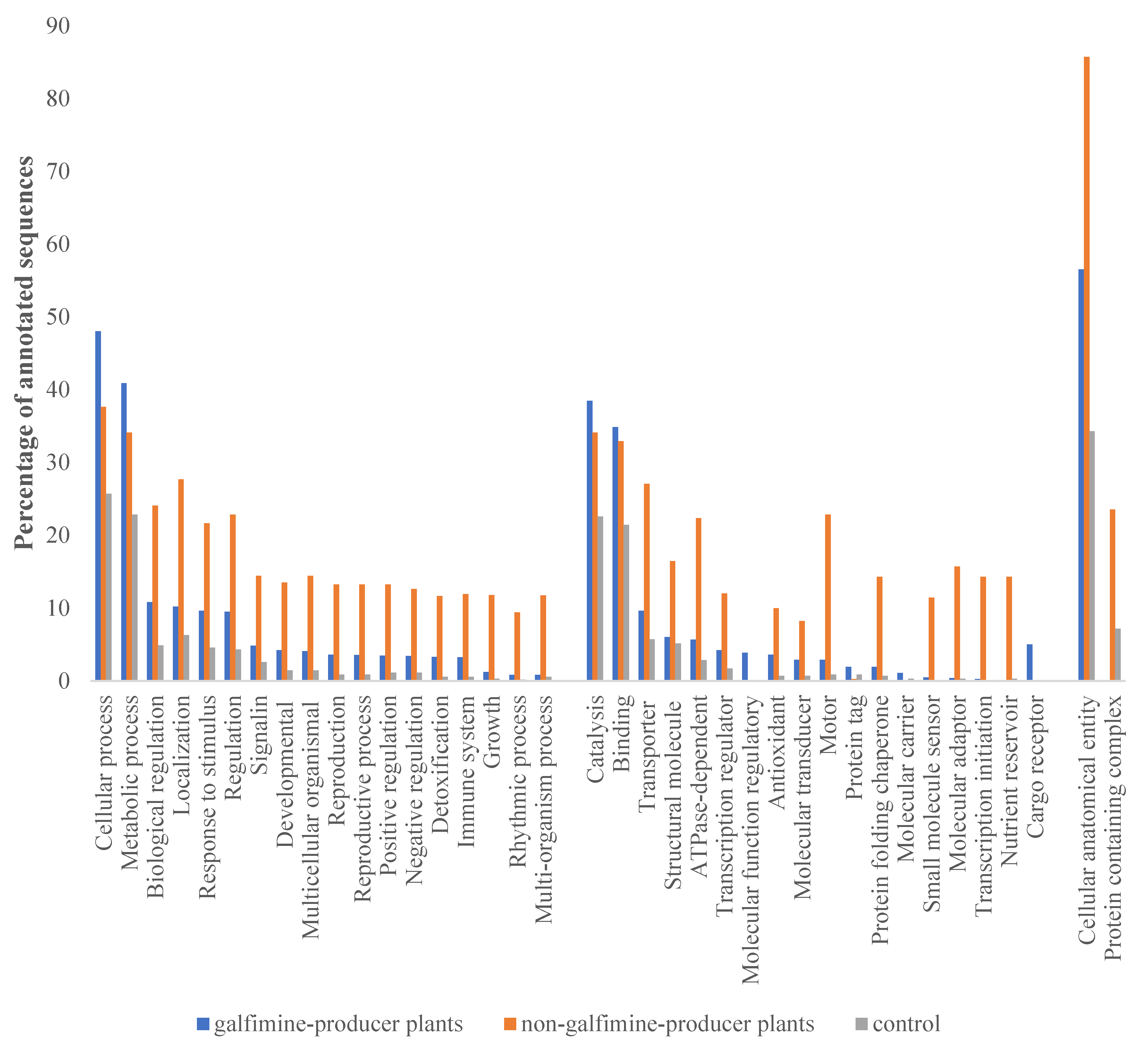
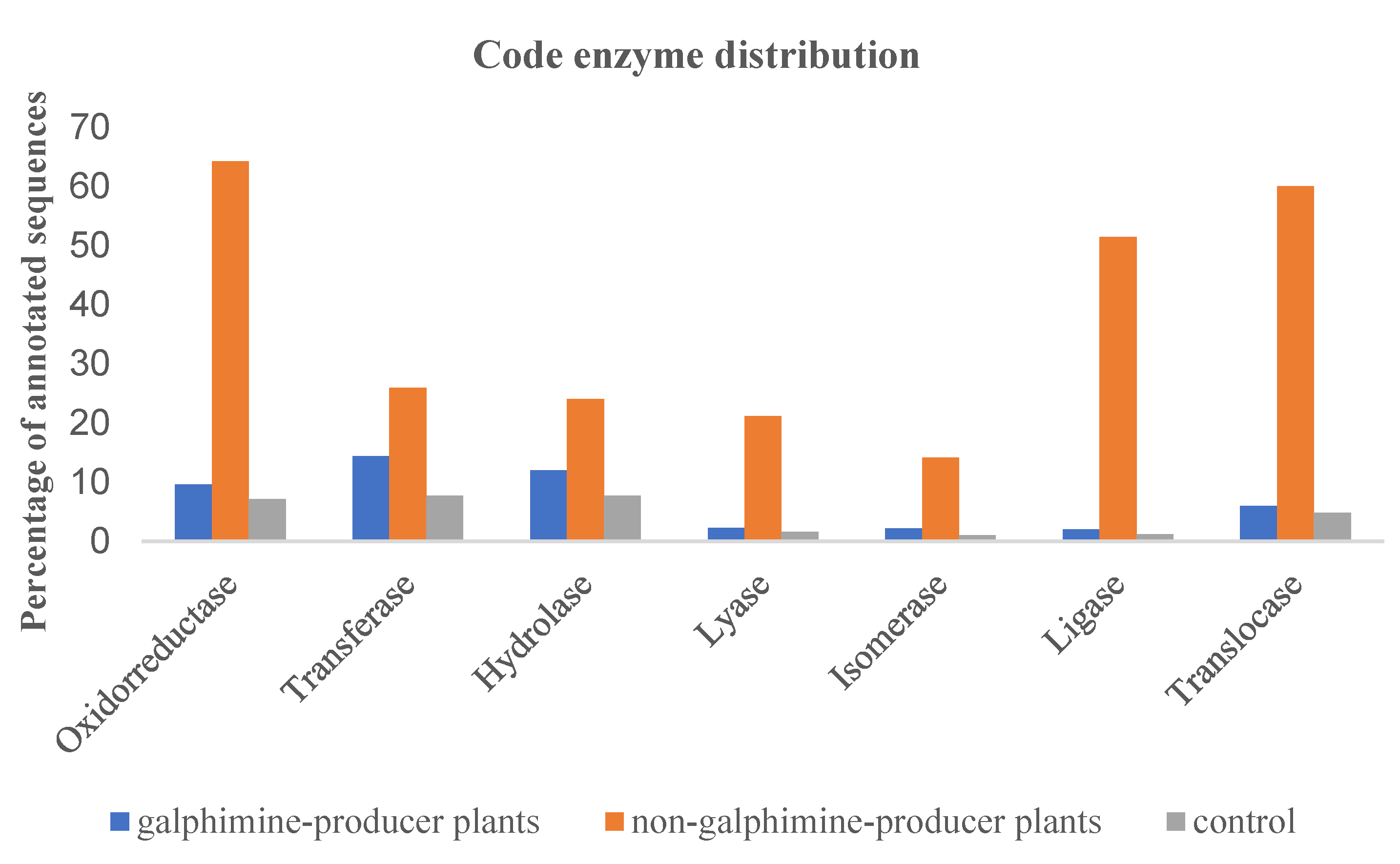
2.3. Transcriptome Functional Annotation
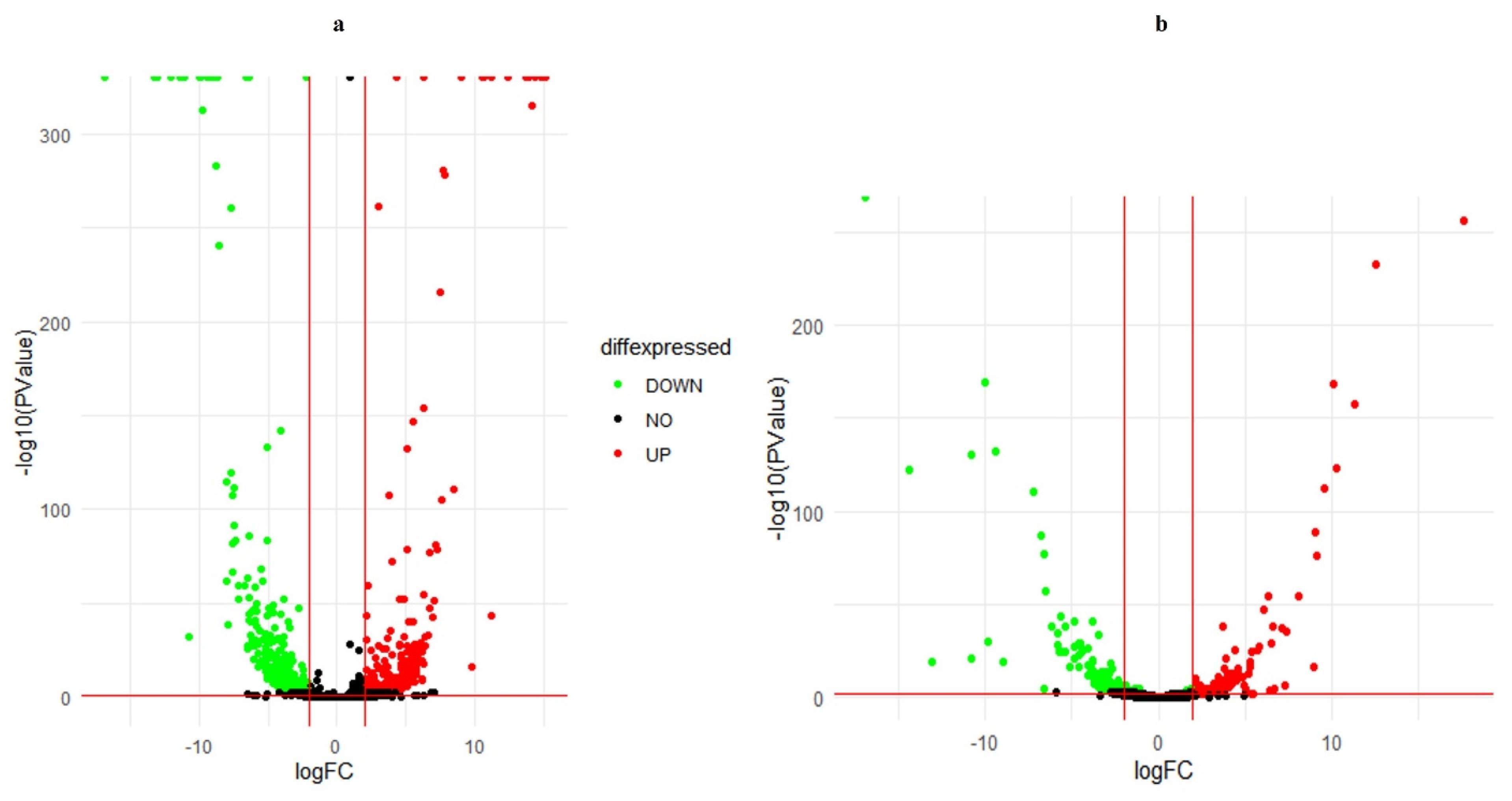
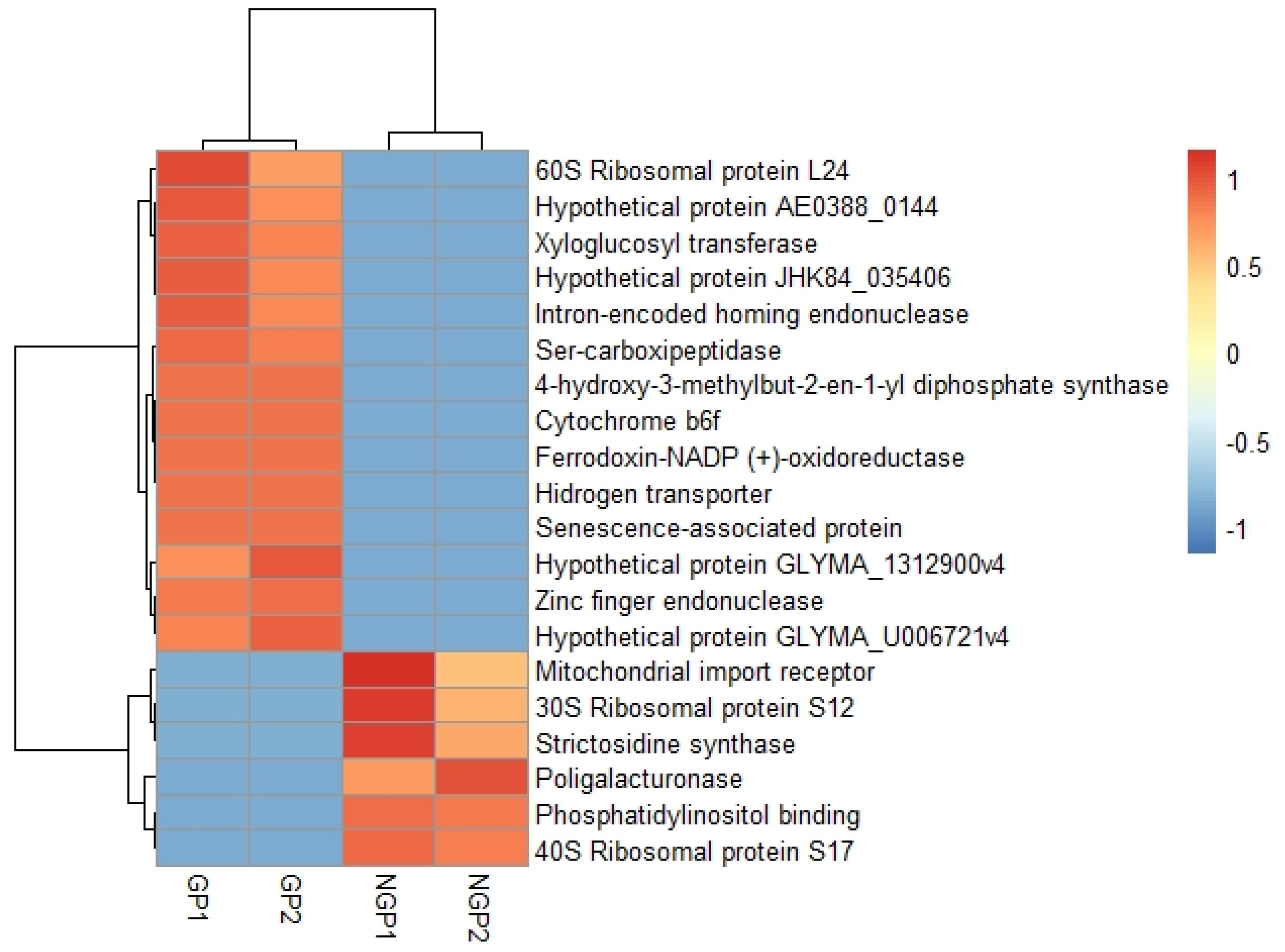
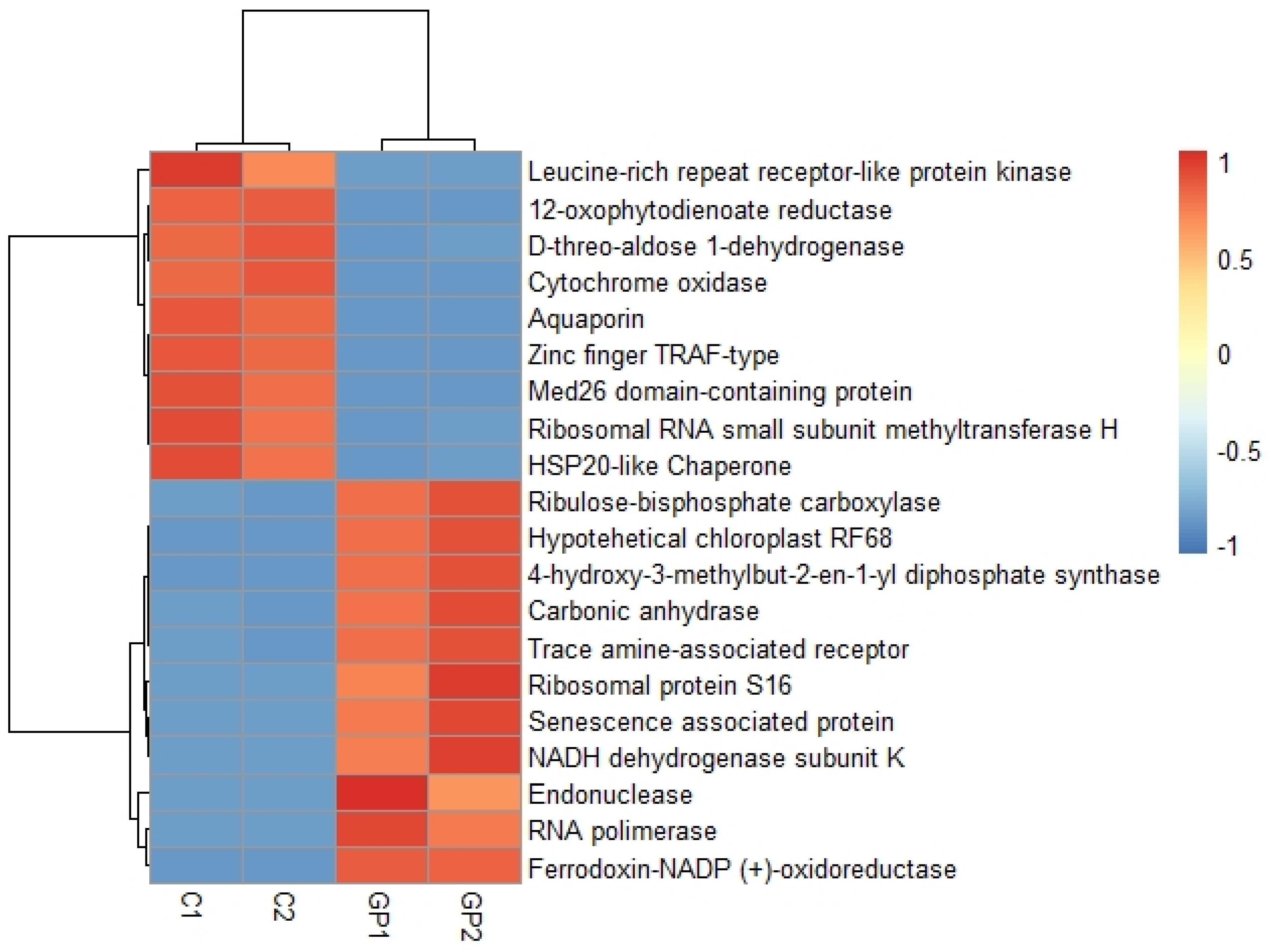
2.4. Differential Expression Analysis
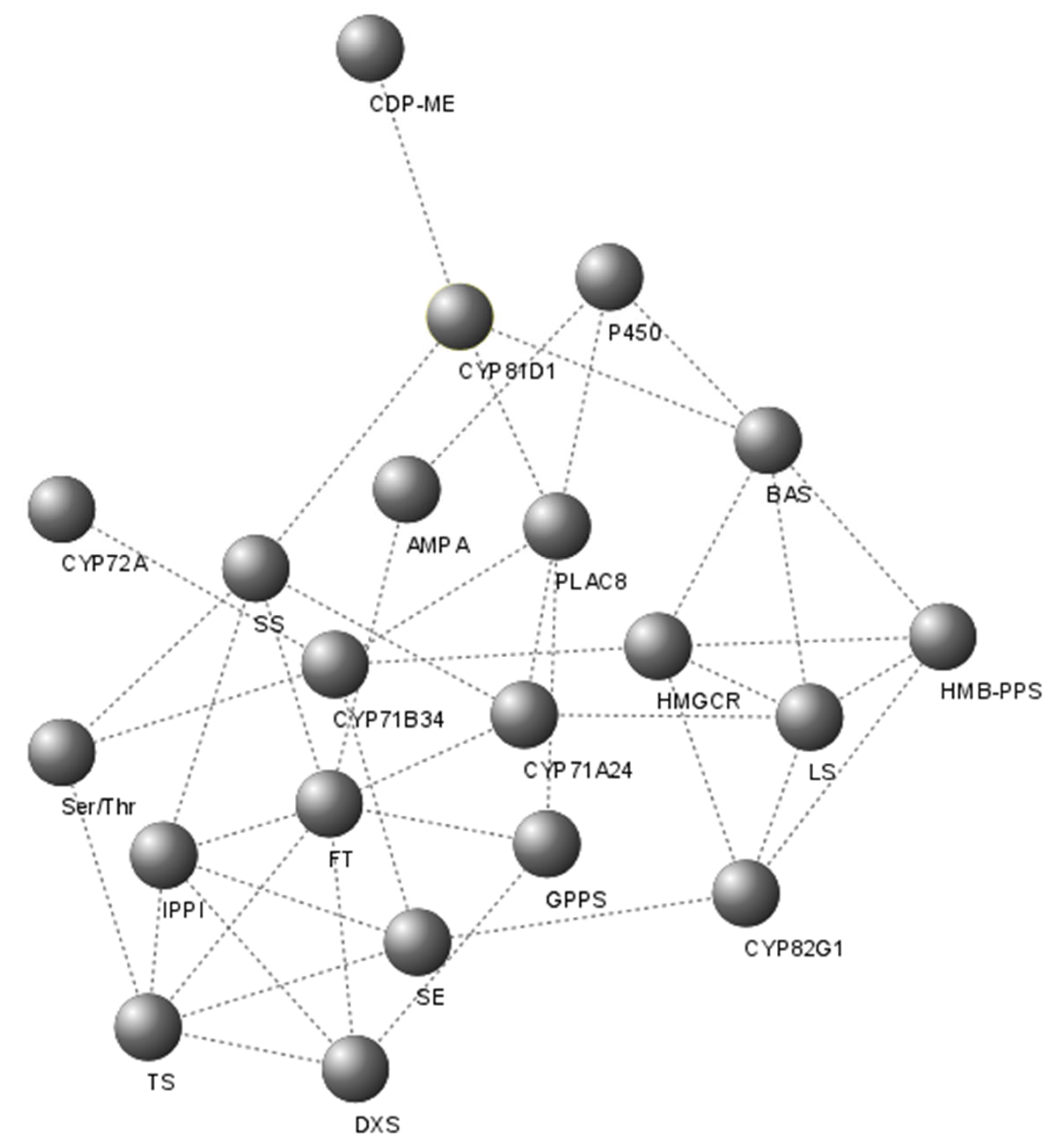
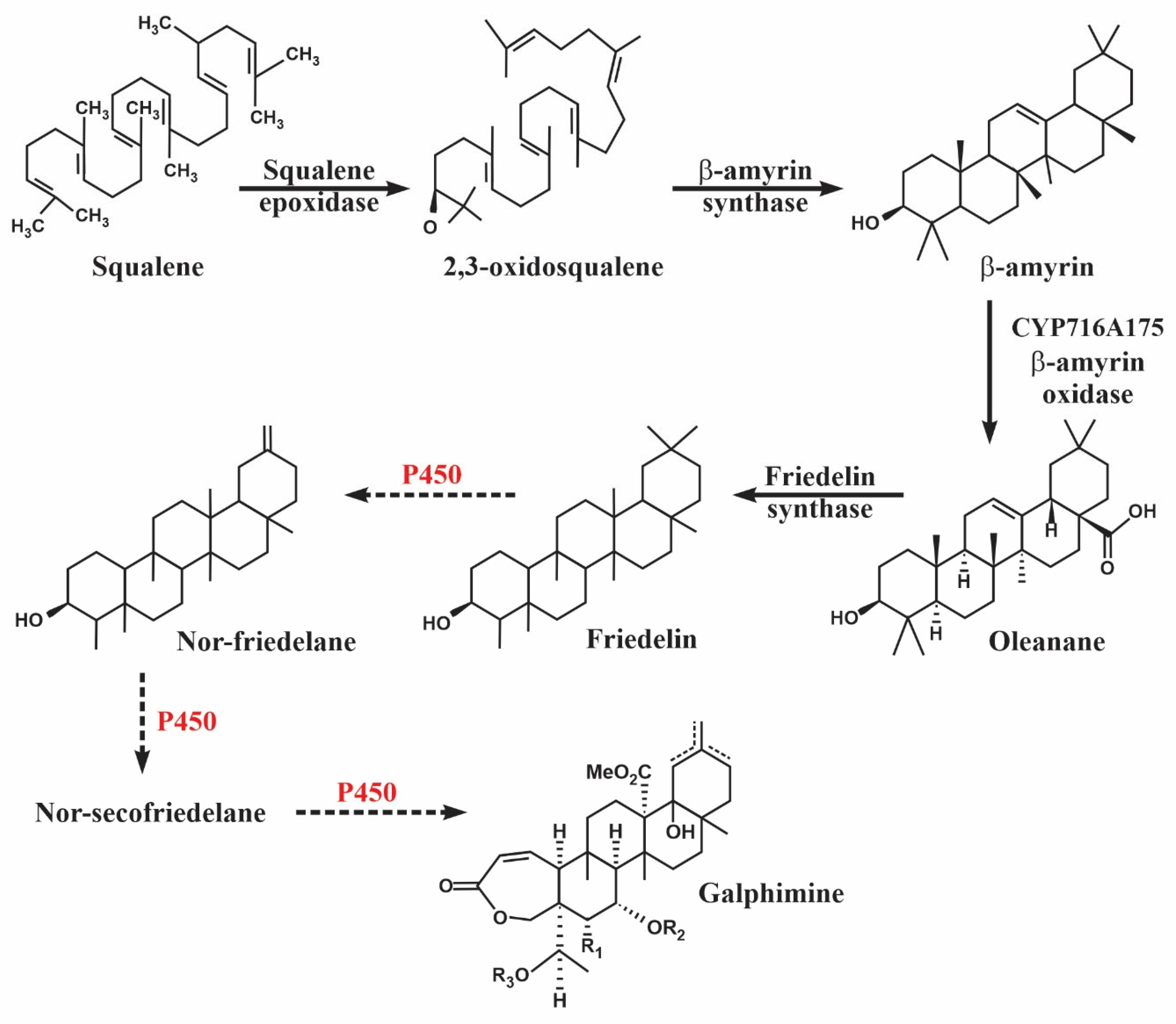
2.5. Identification of Putative Candidate Genes That Encode Enzymes of the Galphimine Synthesis Pathway
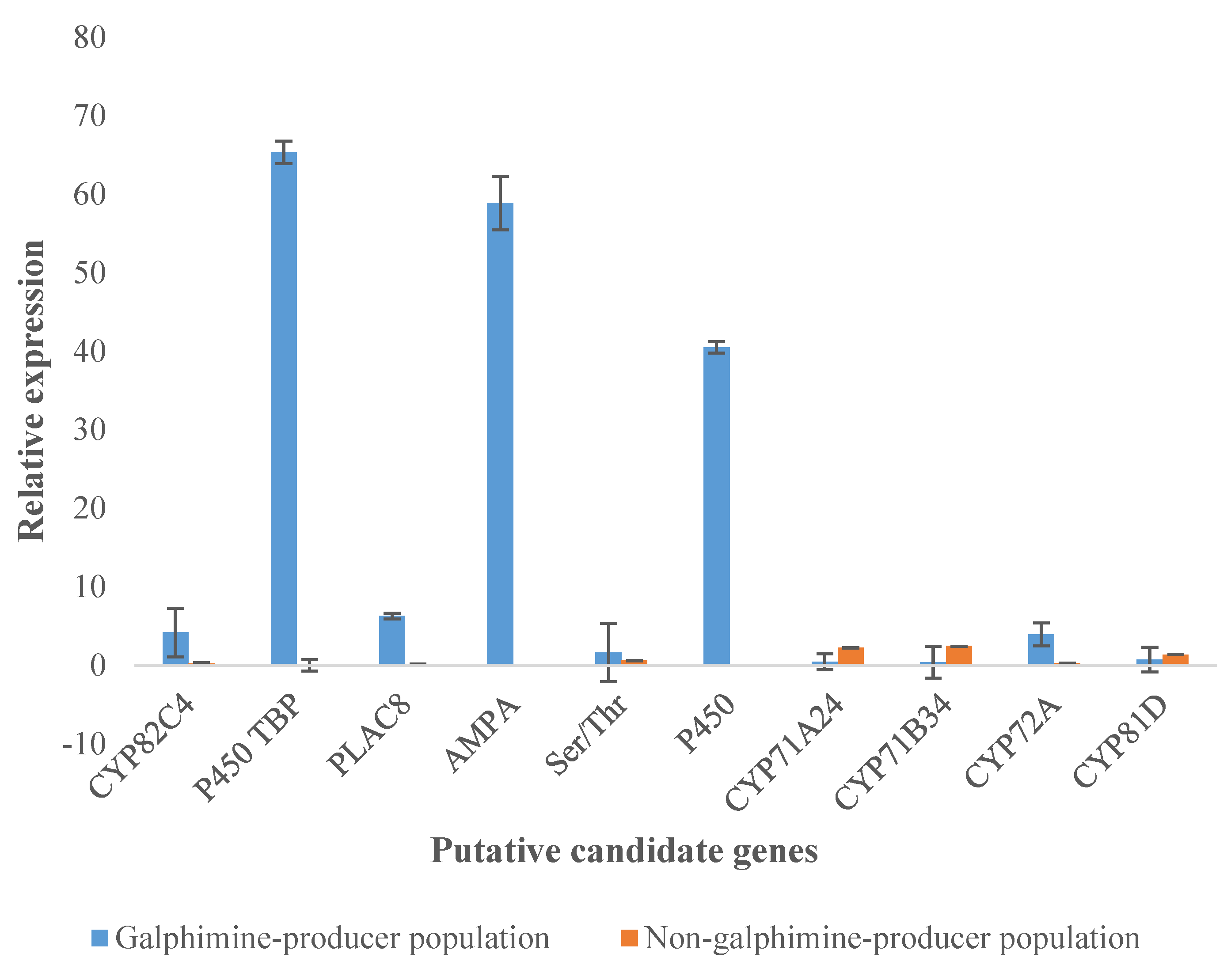
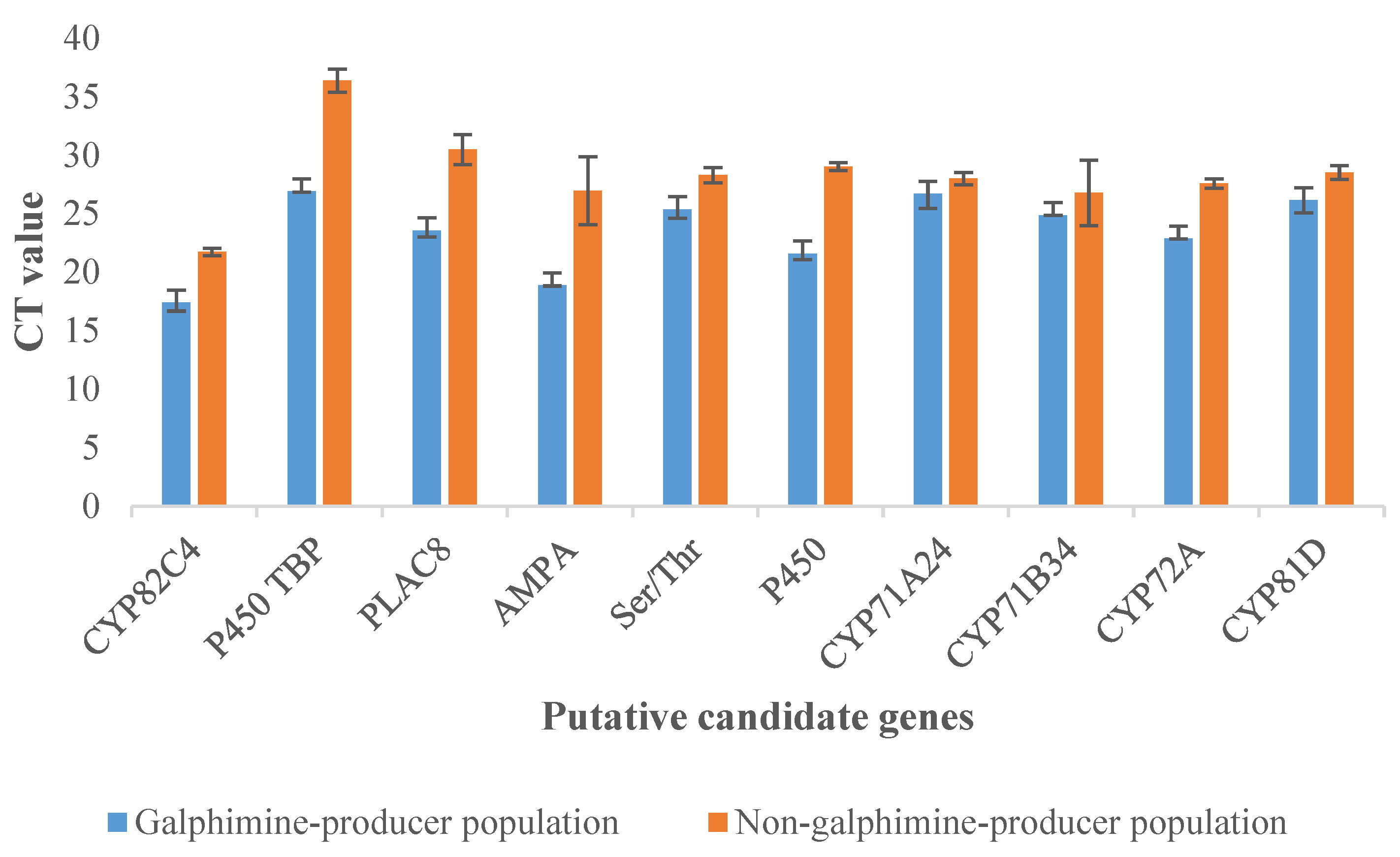
2.6. Validation of Transcriptome Data by qPCR
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. RNA Extraction, Library Preparation, and Transcriptome Sequencing
Reads Quality Control and De Novo Transcriptome Assembly
4.3. Transcriptome Functional Annotation
4.4. Differential Expression Analysis
4.5. Identification of Putative Genes Related to the Galphimines Synthetic Pathway
4.6. Validation of Transcriptome Data by qPCR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tortoriello, J.; Ortega, A.; Herrera-Ruíz, M.; Trujillo, J.; Reyes-Vázquez, C. Galphimine-B modifies electrical activity of ven-tral tegmental area neurons in rats. Planta Med. 1998, 64, 309–313. [Google Scholar] [CrossRef]
- Camacho, M.D.R.; Phillipson, J.D.; Croft, S.L.; Marley, D.; Kirby, G.C.; Warhurst, D.C. Assessment of the Antiprotozoal Activity of Galphimia glauca and the Isolation of New Nor-secofriedelanes and Nor-friedelanes. J. Nat. Prod. 2002, 65, 1457–1461. [Google Scholar] [CrossRef]
- González-Cortazar, M.; Herrera-Ruiz, M.; Zamilpa, A.; Jiménez-Ferrer, E.; Marquina, S.; Álvarez, L.; Tortoriello, J. An-ti-Inflammatory activity and chemical profile of Galphimia glauca. Planta Med. 2014, 80, 90–96. [Google Scholar] [PubMed] [Green Version]
- Campos, M.G.; Toxqui, E.; Tortoriello, J.; Oropeza, M.V.; Ponce, H.; Vargas, M.H.; Montaño, L.M. Galphimia glauca organic fraction antagonizes LTD4-induced contraction in guinea pig airways. J. Ethnopharmacol. 2001, 74, 7–15. [Google Scholar] [CrossRef]
- Dorsch, W.; Wagner, H. New antiasthmatic drugs from traditional medicine? Int. Arch. Allergy Immunol. 1991, 94, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Osuna, L.; Pereda-Miranda, R.; Tortoriello, J.; Villarreal, M.L. Production of the sedative triterpene Galphimine B in Gal-phimia glauca tissue culture. Planta Med. 1999, 65, 149–152. [Google Scholar] [CrossRef]
- Romero-Cerecero, O.; Islas-Garduño, A.L.; Zamilpa, A.; Pérez-García, M.D.; Tortoriello, J. Therapeutic effectiveness of Gal-phimia glauca in young people with social anxiety disorder: A pilot study. Evid. Based Complement. Altern. Med. 2018. [Google Scholar] [CrossRef] [Green Version]
- Taketa, A.T.C.; Lozada-Lechuga, J.; Fragoso-Serrano, M.; Villarreal, M.L.; Pereda-Miranda, R. Isolation of Nor-secofriedelanes from the Sedative Extracts of Galphimia glauca. J. Nat. Prod. 2004, 67, 644–649. [Google Scholar] [CrossRef]
- González-Cortazar, M.; Tortoriello, J.; Alvarez, L. Nor-secofriedelanes as spasmolytics, advances of structure-activity rela-tionships. Planta Med. 2005, 71, 711–716. [Google Scholar] [CrossRef]
- Ortega, A.; Pastor-Palacios, G.; Ortiz-Pastrana, N.; Ávila-Cabezas, E.; Toscano, R.A.; Joseph-Nathan, P.; Morales-Jiménez, J.; Bautista, E. Further galphimines from a new population of Galphimia glauca. Phytochemistry 2019, 169, 112180. [Google Scholar] [CrossRef]
- Náder, B.L.; Cardoso-Taketa, A.T.; Iturriaga, G.; Pereda-Miranda, R.; Villarreal, M.L. Genetic transformation of Galphimia glauca by Agrobacterium rhizogenes and the production of Norfriedelanes. Planta Med. 2004, 70, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Ortíz, A.; Cardoso-Taketa, A.; Rodríguez, M.; Arellano, J.; Hernández, G.; Villarreal, M.L. Transformed cell suspension cul-ture of Galphimia glauca producing sedative Nor-friedelanes. Planta Med. 2010, 76, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Taketa, A.T.; Pereda-Miranda, R.; Choi, Y.H.; Verpoorte, R.; Villarreal, M.L. Metabolic profiling of the Mexican anxiolytic and sedative plant Galphimia glauca using Nuclear Magnetic Resonance spectroscopy and multivariate data analy-sis. Planta Med. 2008, 74, 1295–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Cardoso-Taketa, A.; Choi, Y.H.; Verpoorte, R.; Villarreal, M.L. A comparison on the metabolic profiling of the Mexican anxiolytic and sedative plant Galphimia glauca four years later. J. Ethnopharmacol. 2012, 141, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Folch, J.L.; Cardoso-Taketa, A.; Lorence, A.; Villarreal, M.L. DNA barcoding of the Mexican sedative and anxio-lytic plant Galphimia glauca. J. Ethnopharmacol. 2012, 144, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gesto-Borroto, R.; Cardoso-Taketa, A.; Yactayo-Chang, J.P.; Medina-Jiménez, K.; Hornung-Leoni, C.; Lorence, A.; Villarreal, M.L. DNA barcoding and TLC as tools to properly identify natural populations of the Mexican medicinal species Galphimia glauca Cav. PLoS ONE 2019, 14, e0217313. [Google Scholar] [CrossRef] [Green Version]
- Toscano, R.A.; Maldonado, E.; Lozoya, X.; Tortoriello, J.; Ortega, A.; Gaviño, R. Structure of galphimine B. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1993, 49, 774–776. [Google Scholar] [CrossRef]
- Ma, C.H.; Gao, Z.J.; Zhang, J.J.; Zhang, W.; Shao, J.H.; Hai, M.R.; Chen, J.W.; Yang, S.C.; Zhang, G.H. Candidate genes in-volved in the biosynthesis of triterpenoid saponins in Platycodon grandiflorum identified by transcriptome analysis. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, G.; Sandeep; Garg, A.; Misra, R.C.; Chanotiya, C.S.; Ghosh, S. Transcriptome analysis and functional characterization of oxidosqualene cyclases of the arjuna triterpene saponin pathway. Plant Sci. 2019, 292, 110382. [Google Scholar] [CrossRef]
- Lertphadungkit, P.; Qiao, X.; Sirikantaramas, S.; Satitpatipan, V.; Ye, M.; Bunsupa, S. De novo transcriptome analysis and identification of candidate genes associated with triterpenoid biosynthesis in Trichosanthes cucumerina L. Plant Cell Rep. 2021, 40, 1845–1858. [Google Scholar] [CrossRef]
- Rios, M.Y.; Ortega, A.; Domínguez, B.; Déciga, M.; de la Rosa, V. Glaucacetalin E and galphimidin B from Galphimia glauca and their anxiolytic activity. J. Ethnopharmacol. 2020, 259, 112939. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C. Revision of Galphimia (Malpighiaceae). Contrib. Univ. Mich. Herb. 2007, 25, 1–82. [Google Scholar]
- Ghosh, S. Triterpene Structural Diversification by Plant Cytochrome P450 Enzymes. Front. Plant Sci. 2017, 8, 1886. [Google Scholar] [CrossRef]
- Bathe, U.; Tissier, A. Cytochrome P450 enzymes. Phytochemistry 2019, 161, 149–162. [Google Scholar] [CrossRef]
- Geisler, K.; Hughes, R.K.; Sainsbury, F.; Lomonossoff, G.P.; Rejzek, M.; Fairhurst, S.; Olsen, C.E.; Motawia, M.S.; Melton, R.E.; Hemmings, A.M.; et al. Biochemical analysis of a multifunctional cytochrome P450 (CYP51) enzyme required for synthesis of antimicrobial triterpenes in plants. Proc. Natl. Acad. Sci. USA 2013, 110, 3360–3367. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Hu, T.; Gao, L.; Su, P.; Zhang, Y.; Zhao, Y.; Chen, S.; Tu, L.; Song, Y.; Wang, X.; et al. Friedelane-type triterpene cyclase in celastrol biosynthesis from Tripterygium wilfordii and its application for triterpenes biosynthesis in yeast. New Phytol. 2019, 223, 722–735. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, H.; Tong, X.; Liu, Z.; Zhang, X.; Li, D.; Jiang, X.; Yu, X. Identification and analysis of CYP450 and UGT super-gene family members from the transcriptome of Aralia elata (Miq.) seem to reveal candidate genes for triterpenoid saponin biosynthesis. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Souza-Moreira, T.M.; Alves, T.B.; Pinheiro, K.; Felippe, L.G.; De Lima, G.M.A.; Watanabe, T.F.; Barbosa, C.C.; Santos, V.A.F.F.M.; Lopes, N.P.; Valentini, S.R.; et al. Friedelin Synthase from Maytenus ilicifolia: Leucine 482 Plays an Essential Role in the Production of the Most Rearranged Pentacyclic Triterpene. Sci. Rep. 2016, 6, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. In Biotechnology of Isoprenoids. Advances in Biochemical Engineering/Biotechnology, 1st ed.; Chrader, J., Bohlmann, J., Eds.; Springer: Luxemburgo, 2015; Volume 148, pp. 63–106. [Google Scholar]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene synthases from Cannabis sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Badieyan, S.; Bevan, D.R.; Herde, M.; Gatz, C.; Tholl, D. Herbivore-induced, and floral homoterpene volatiles are biosynthesized by a single P450 enzyme (CYP82G1) in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 21205–21210. [Google Scholar] [CrossRef] [Green Version]
- Hartl, C.L.; Ramaswami, G.; Pembroke, W.G.; Muller, S.; Pintacuda, G.; Saha, A.; Parsana, P.; Battle, A.; Lage, K.; Geschwind, D.H. Coexpression network architecture reveals the brain-wide and multiregional basis of disease susceptibility. Nat. Neurosci. 2021, 24, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Itoh, D.; Kawano, A.K.; Nabeta, K. Biosynthesis of Chloroplastidic and Extrachloroplastidic Terpenoids in Liverwort Cultured Cells: 13C Serine as a Probe of Terpene Biosynthesis via Mevalonate and Non-mevalonate Pathways. J. Nat. Prod. 2003, 66, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Chu, L.; Shu, W.; He, X.; Wang, L.; Lu, M. Selection and validation of reference genes for quantitative expression analysis of miRNAs and mRNAs in Poplar. Plant Methods 2019, 15, 35. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample and Replica | Number of Transcripts |
|---|---|
| Leaves of galphimine producing-population | 9023 |
| Leaves of galphimine producing population, replica | 8915 |
| Leaves of non-galphimine producing-population | 8667 |
| Leaves of non-galphimine producing-population, replica | 8896 |
| Roots of galphimine producing-population | 5004 |
| Roots of galphimine producing population, replica | 4667 |
| Galphimine-Producer Population | Non-Galphimine-Producer Population | Control | ||||
|---|---|---|---|---|---|---|
| Annotated Transcripts | Number of Annotations | Gen Counts | Number of Annotations | Gen Counts | Number of Annotations | Gen Counts |
| 3-hydroxy-3-methylglutaryl-CoA reductase | 2 | 13 | 1 | 8 | 1 | 6 |
| 1-deoxy-D-xylulose-5-phosphate synthase | 4 | 15 | 4 | 29 | - | - |
| 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase | 4 | 18 | 1 | 10 | - | - |
| 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase | 5 | 19 | 5 | 16 | - | - |
| isopentenyl diphosphate isomerase | 1 | 52 | 1 | 17 | - | - |
| farnesyl transferase | 1 | 6 | 3 | 4 | - | - |
| geranyl diphosphate synthase | 1 | 14 | 1 | 4 | - | - |
| terpene synthase | 1 | 347 | 3 | 253 | 5 | 78 |
| squalene synthase | 1 | 7 | - | - | - | - |
| squalene epoxidase | 1 | 16 | - | - | - | - |
| beta-amyrin synthase | 2 | 23 | 4 | 14 | - | - |
| lanosterol synthase | 7 | 18 | 6 | 23 | - | - |
| P450 | 2 | 304.444 | - | - | 1 | 4 |
| P450 TBP | 19 | 417.183 | - | - | - | - |
| CYP71A24 | 1 | 5 | 2 | 3 | - | - |
| CYP71A26 | - | - | 1 | 14 | - | - |
| CYP71B10 | - | - | 1 | 12 | - | - |
| CYP71B34 | 1 | 9 | - | - | - | - |
| CYP71D11 | - | - | - | - | 1 | 15 |
| CYP72A | 2 | 10 | - | - | - | - |
| CYP72A15 | - | - | 1 | 7 | - | - |
| CYP81D1 | 1 | 18 | - | - | - | - |
| CYP82C4 | 1 | 231 | - | - | - | - |
| CYP82G1 | 1 | 41 | 1 | 129 | - | - |
| PLAC8 | 2 | 12 | - | - | - | - |
| 5’-AMP-activated protein kinase | 1 | 30 | - | - | - | - |
| serine/threonine-protein kinase AtPK2/AtPK19 | 1 | 6 | - | - | - | - |
| Gene Description | Gene Symbol | Primer Sequence 5′-3′ | PCR Product Length (bp) |
|---|---|---|---|
| CYP82C4 | CYP82C4 | F 5′TCATTTGGTGGGCTAAAAGC 3′ R 5′ACGAGTCCGTTAATGGTTGC 3′ | 191 |
| P450 TBP | P450 TBP | F 5′AGCTTCGTCGCAAGTGAAAT 3′ R 5′TAGGACGCCTGCGTTATCTT 3′ | 189 |
| PLAC8 | PLAC8 | F 5′ACTGTTGATAACCCCGATGG 3′ R 5′TCTCAGGAAGTCCGAACTGG 3′ | 170 |
| 5′-AMP-activated protein kinase | AMPA | F 5′CCCAAGGAAAAGGTTTCACA 3′ R 5′GGCAAAAGCAATGGCTAAGA 3′ | 229 |
| serine/threonine-protein kinase AtPK2/AtPK19 | Ser/Thr | F 5′TTTCAACTGGACCACAAAGG 3′ R 5′CGTTCGGGAAAGTCTACCAA 3′ | 164 |
| P450 | P450 | F 5′CCTTGAGGTCATGGCTGAAT 3′ R 5′GCTTCTCTCCAAAGGCACAC 3′ | 232 |
| CYP71A24 | CYP71A24 | F 5′CAAACCGGCCTAAATCAAAG 3′ R 5′GCTCTTGTTCATAACTTTCTCAATC 3′ | 200 |
| CYP71B34 | CYP71B34 | F 5′CCGTGAGAGAGGCCATTAAC 3′ R 5′GTGCGAAAGATTTGCGTTCT 3′ | 160 |
| CYP72A | CYP72A | F 5′TTGTTGGCTTTTCGAGGAAT 3′ R 5′AAGCATCAGGAGTGGCAAAC 3′ | 167 |
| CYP81D1 | CYP81D1 | F 5′TCGGAGGATTGGACTACGAC 3′ R 5′TTCCGCCATAACATTTCTCC 3′ | 219 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iglesias, D.; Donato Capote, M.d.; Méndez Tenorio, A.; Valdivia, A.V.; Gutiérrez-García, C.; Paul, S.; Iqbal, H.M.N.; Villarreal, M.L.; Sharma, A. Identification of Putative Candidate Genes from Galphimia spp. Encoding Enzymes of the Galphimines Triterpenoids Synthesis Pathway with Anxiolytic and Sedative Effects. Plants 2022, 11, 1879. https://doi.org/10.3390/plants11141879
Iglesias D, Donato Capote Md, Méndez Tenorio A, Valdivia AV, Gutiérrez-García C, Paul S, Iqbal HMN, Villarreal ML, Sharma A. Identification of Putative Candidate Genes from Galphimia spp. Encoding Enzymes of the Galphimines Triterpenoids Synthesis Pathway with Anxiolytic and Sedative Effects. Plants. 2022; 11(14):1879. https://doi.org/10.3390/plants11141879
Chicago/Turabian StyleIglesias, Dianella, Marcos de Donato Capote, Alfonso Méndez Tenorio, Ana Victoria Valdivia, Claudia Gutiérrez-García, Sujay Paul, Hafiz M. N. Iqbal, María Luisa Villarreal, and Ashutosh Sharma. 2022. "Identification of Putative Candidate Genes from Galphimia spp. Encoding Enzymes of the Galphimines Triterpenoids Synthesis Pathway with Anxiolytic and Sedative Effects" Plants 11, no. 14: 1879. https://doi.org/10.3390/plants11141879
APA StyleIglesias, D., Donato Capote, M. d., Méndez Tenorio, A., Valdivia, A. V., Gutiérrez-García, C., Paul, S., Iqbal, H. M. N., Villarreal, M. L., & Sharma, A. (2022). Identification of Putative Candidate Genes from Galphimia spp. Encoding Enzymes of the Galphimines Triterpenoids Synthesis Pathway with Anxiolytic and Sedative Effects. Plants, 11(14), 1879. https://doi.org/10.3390/plants11141879