

Glioma-Associated Sialoglycans Drive the Immune Suppressive Phenotype and Function of Myeloid Cells
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Glioma Cell Culture and Sialic Acid Inhibitor Treatment
2.2. Monocyte Isolation and Co-Culture Experiments
2.3. Siglec Activation in Monocytes
2.4. Flow Cytometry and Cytokine ELISA
2.5. Statistical Analysis
3. Results
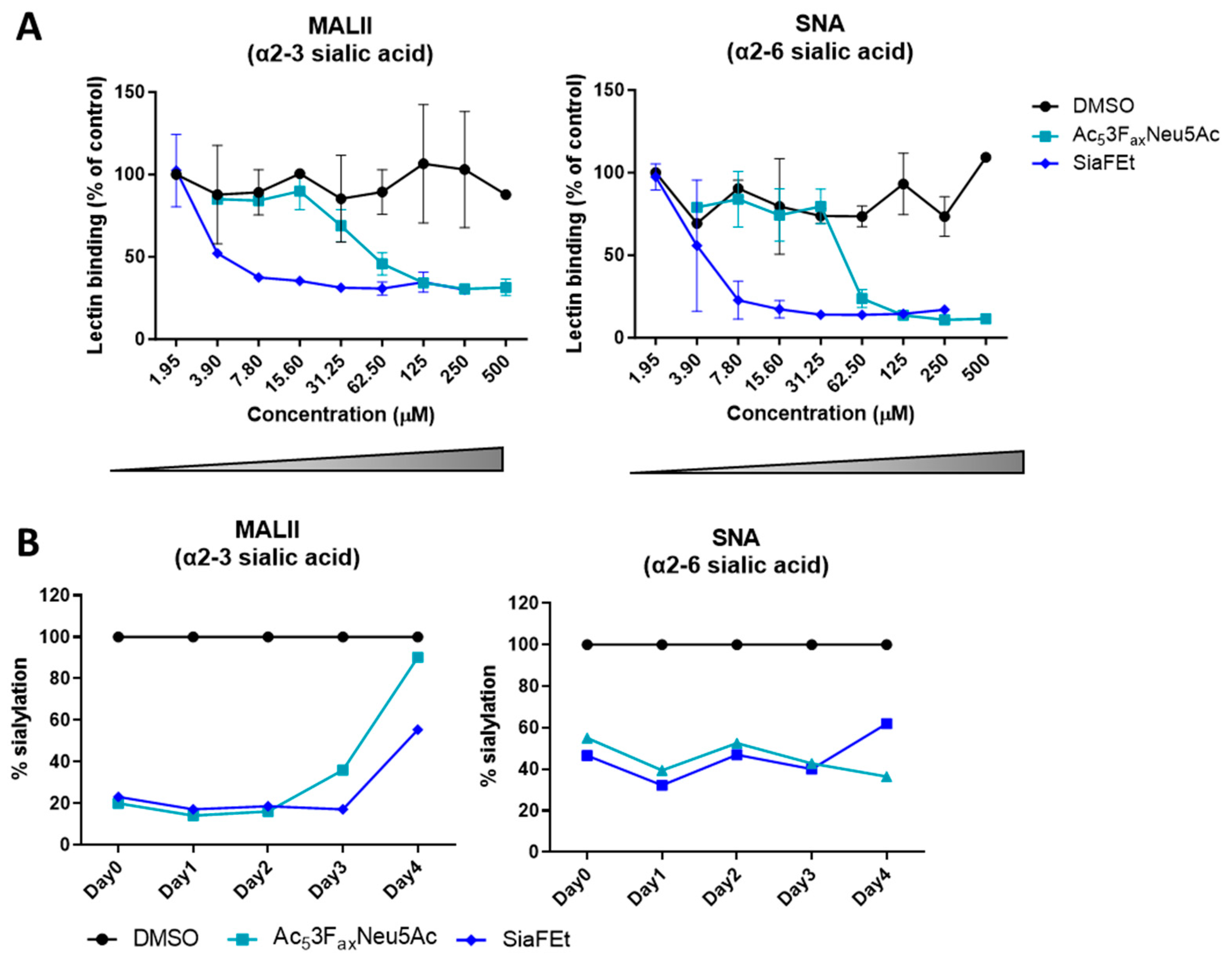
3.1. Sialic Acid Inhibition Results in Long-Lasting Reduction in Sialylation in the Glioblastoma T98G Cell Line
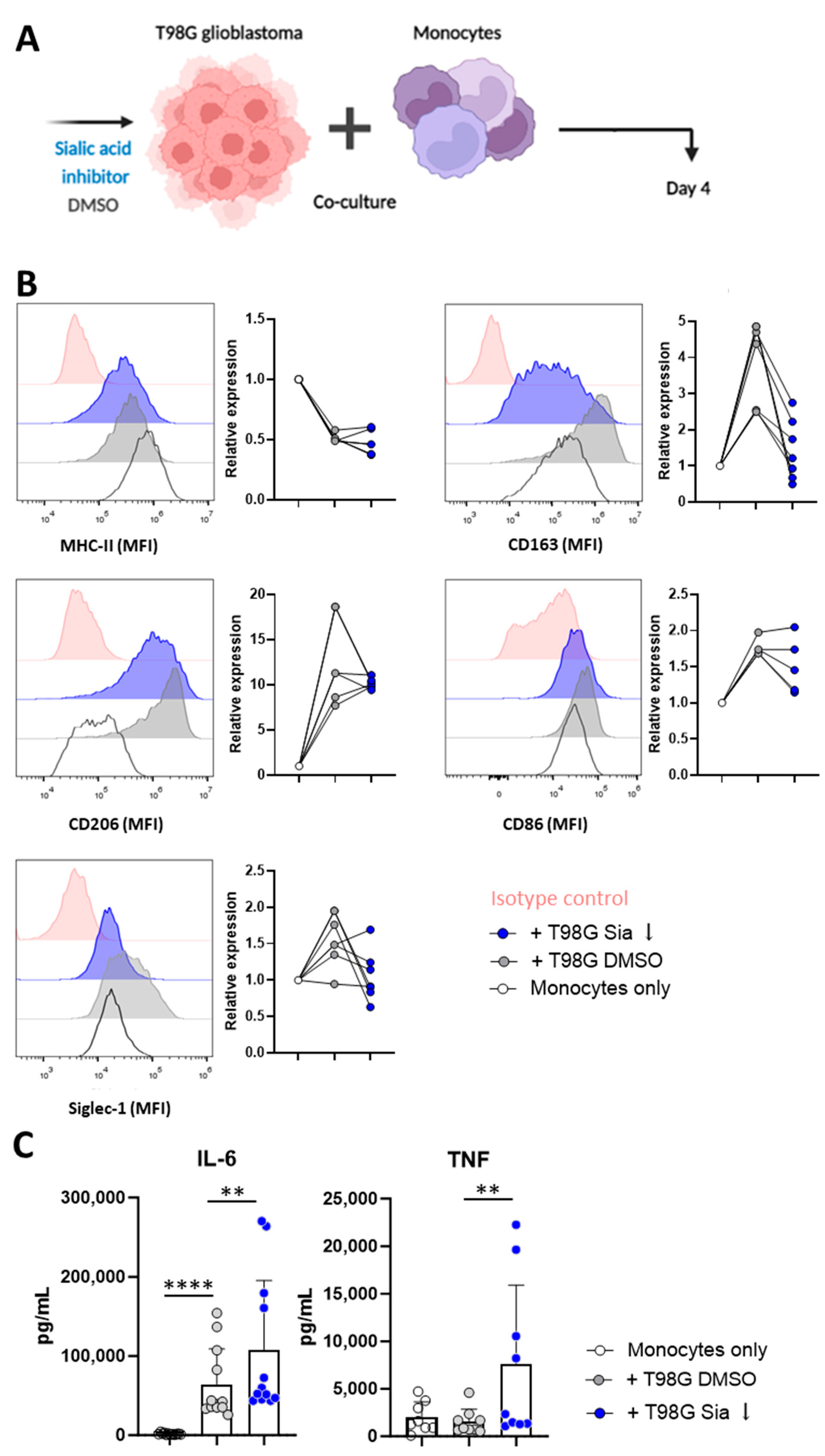
3.2. Glioblastoma Cells Induce Sialic Acid Dependent Immune Suppressive Phenotypes in Monocytes
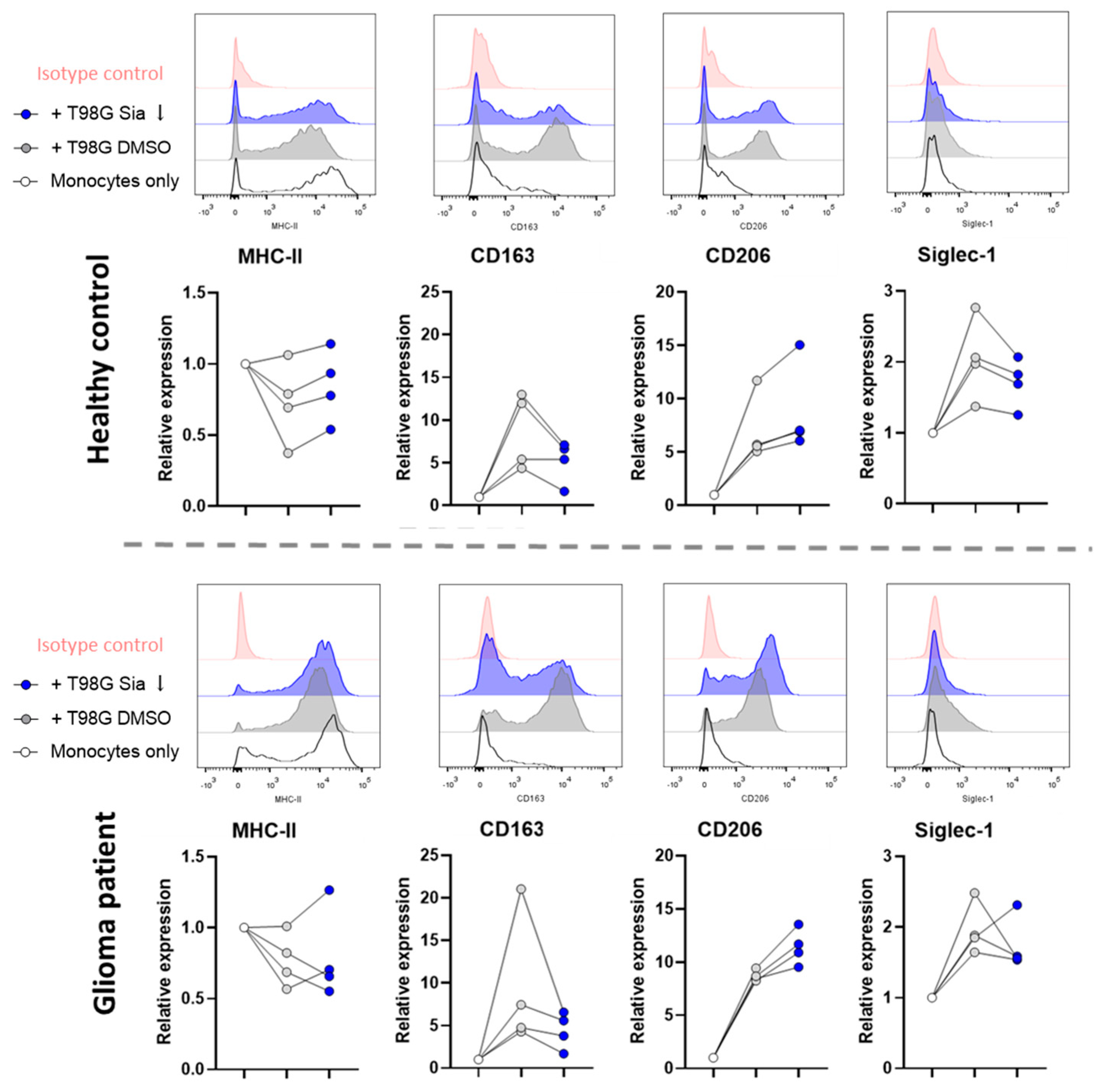
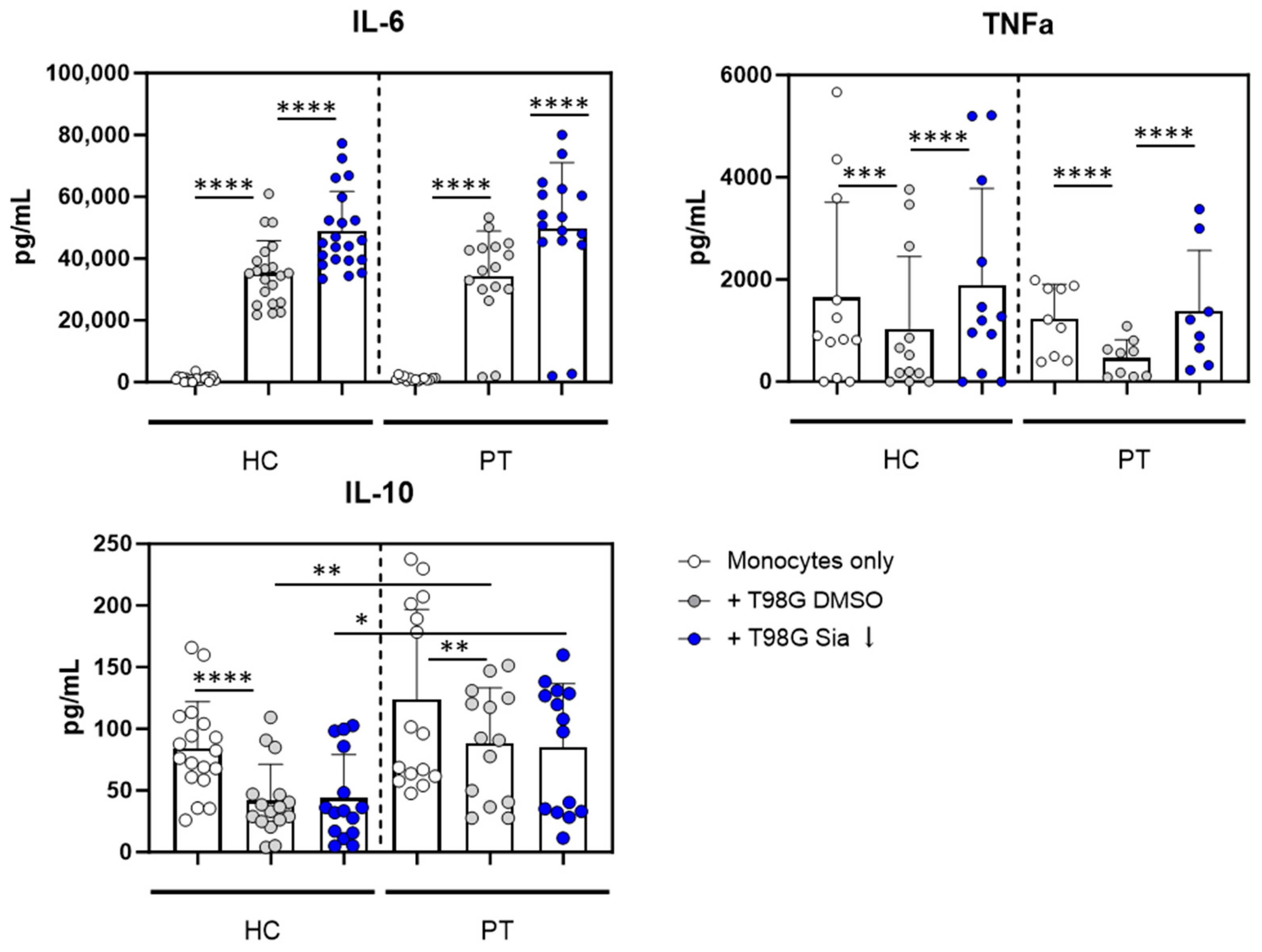
3.3. Glioblastoma Cells Induce Sialic Acid Dependent Immune Suppressive Phenotypes in Glioma Patient Derived Monocytes
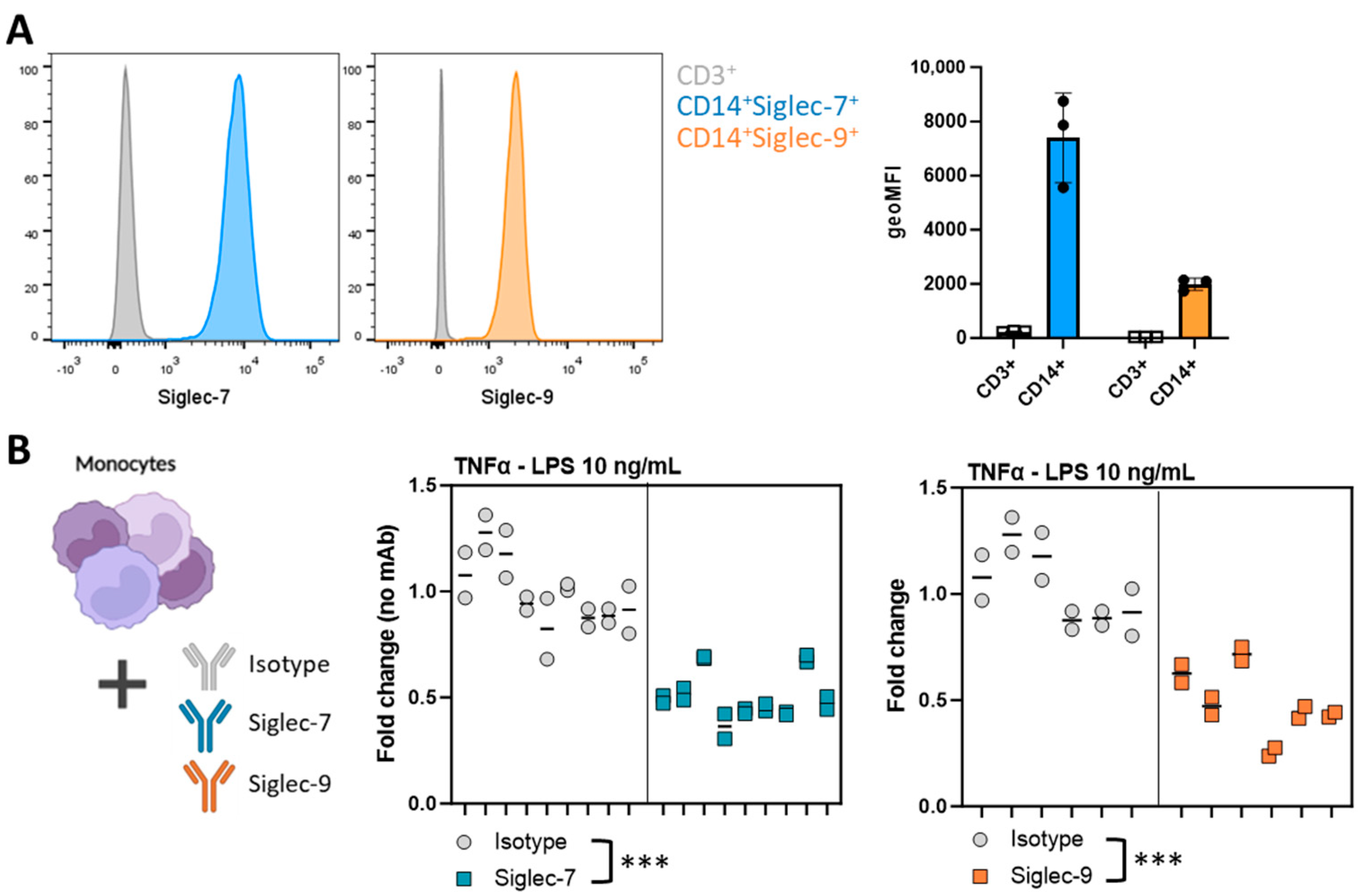
3.4. Siglec Activation Validates the Involvement of Siglec Receptors in the T98G-Induced Immunosuppressive Myeloid Phenotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akhavan, O.; Ghaderi, E. Graphene Nanomesh Promises Extremely Efficient In Vivo Photothermal Therapy. Small 2013, 9, 3593–3601. [Google Scholar] [CrossRef] [PubMed]
- McGranahan, T.; Li, G.; Nagpal, S. History and Current State of Immunotherapy in Glioma and Brain Metastasis. Ther. Adv. Med. Oncol. 2017, 9, 347. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Roth, P.; Preusser, M.; Wick, W.; Reardon, D.A.; Platten, M.; Sampson, J.H. Vaccine-Based Immunotherapeutic Approaches to Gliomas and Beyond. Nat. Rev. Neurol. 2017, 13, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Gielen, P.R.; Schulte, B.M.; Kers-Rebel, E.D.; Verrijp, K.; Petersen-Baltussen, H.M.J.M.; Ter Laan, M.; Wesseling, P.; Adema, G.J. Increase in Both CD14-Positive and CD15-Positive Myeloid-Derived Suppressor Cell Subpopulations in the Blood of Patients with Glioma but Predominance of CD15-Positive Myeloid-Derived Suppressor Cells in Glioma Tissue. J. Neuropathol. Exp. Neurol. 2015, 74, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Raychaudhuri, B.; Ireland, P.R.J.; Ko, J.; Rini, B.; Borden, E.C.; Garcia, J.; Vogelbaum, M.A.; Finke, J. Myeloid-Derived Suppressor Cell Accumulation and Function in Patients with Newly Diagnosed Glioblastoma. Neuro-Oncology 2011, 13, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Gielen, P.R.; Schulte, B.M.; Kers-Rebel, E.D.; Verrijp, K.; Bossman, S.A.J.F.H.; Ter Laan, M.; Wesseling, P.; Adema, G.J. Elevated Levels of Polymorphonuclear Myeloid-Derived Suppressor Cells in Patients with Glioblastoma Highly Express S100A8/9 and Arginase and Suppress T Cell Function. Neuro-Oncology 2016, 18, 1253–1264. [Google Scholar] [CrossRef]
- van de Wall, S.; Santegoets, K.C.M.; van Houtum, E.J.H.; Büll, C.; Adema, G.J. Sialoglycans and Siglecs Can Shape the Tumor Immune Microenvironment. Trends Immunol. 2020, 41, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Stanczak, M.A.; Läubli, H. Siglec Receptors as New Immune Checkpoints in Cancer. Mol. Asp. Med. 2022, 90, 101112. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.Y.; Qi, L.L.; Kang, F.B.; Wang, L. The Intriguing Roles of Siglec Family Members in the Tumor Microenvironment. Biomark. Res. 2022, 10, 22. [Google Scholar] [CrossRef]
- Chen, X.; Eksioglu, E.A.; Zhou, J.; Zhang, L.; Djeu, J.; Fortenbery, N.; Epling-Burnette, P.; Van Bijnen, S.; Dolstra, H.; Cannon, J.; et al. Induction of Myelodysplasia by Myeloid-Derived Suppressor Cells. J. Clin. Investig. 2013, 123, 4595–4611. [Google Scholar] [CrossRef]
- Schneider, F.; Kemmner, W.; Haensch, W.; Franke, G.; Gretschel, S.; Karsten, U.; Schlag, P.M. Overexpression of Sialyltransferase CMP-Sialic Acid: Galβ1,3GalNAc-R A6-Sialyltransferase Is Related to Poor Patient Survival in Human Colorectal Carcinomas. Cancer Res. 2001, 61, 4605–4611. [Google Scholar] [PubMed]
- Yeh, S.C.; Wang, P.Y.; Lou, Y.W.; Khoo, K.H.; Hsiao, M.; Hsu, T.L.; Wong, C.H. Glycolipid GD3 and GD3 Synthase Are Key Drivers for Glioblastoma Stem Cells and Tumorigenicity. Proc. Natl. Acad. Sci. USA 2016, 113, 5592–5597. [Google Scholar] [CrossRef] [PubMed]
- van Houtum, E.J.H.; Büll, C.; Cornelissen, L.A.M.; Adema, G.J. Siglec Signaling in the Tumor Microenvironment. Front. Immunol. 2021, 12, 790317. [Google Scholar] [CrossRef]
- Varki, A. Glycan-Based Interactions Involving Vertebrate Sialic-Acid-Recognizing Proteins. Nature 2007, 446, 1023–1029. [Google Scholar] [CrossRef]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.D.; Klausing, S.; Hillier, M.; Maher, J.; Noll, T.; Crocker, P.R.; et al. The Mucin MUC1 Modulates the Tumor Immunological Microenvironment through Engagement of the Lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Santegoets, K.C.M.; Gielen, P.R.; Büll, C.; Schulte, B.M.; Kers-Rebel, E.D.; Küsters, B.; Bossman, S.A.J.F.H.; ter Laan, M.; Wesseling, P.; Adema, G.J. Expression Profiling of Immune Inhibitory Siglecs and Their Ligands in Patients with Glioma. Cancer Immunol. Immunother. 2019, 68, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Schmassmann, P.; Roux, J.; Buck, A.; Tatari, N.; Hogan, S.; Wang, J.; Rodrigues Mantuano, N.; Wieboldt, R.; Lee, S.; Snijder, B.; et al. Targeting the Siglec-Sialic Acid Axis Promotes Antitumor Immune Responses in Preclinical Models of Glioblastoma. Sci. Transl. Med. 2023, 15, eadf5302. [Google Scholar] [CrossRef]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Azadi, P.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global Metabolic Inhibitors of Sialyl- and Fucosyltransferases Remodel the Glycome. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef]
- Büll, C.; Boltje, T.J.; Van Dinther, E.A.W.; Peters, T.; De Graaf, A.M.A.; Leusen, J.H.W.; Kreutz, M.; Figdor, C.G.; Den Brok, M.H.; Adema, G.J. Targeted Delivery of a Sialic Acid-Blocking Glycomimetic to Cancer Cells Inhibits Metastatic Spread. ACS Nano 2015, 9, 733–745. [Google Scholar] [CrossRef]
- Heise, T.; Pijnenborg, J.F.A.; Büll, C.; Van Hilten, N.; Kers-Rebel, E.D.; Balneger, N.; Elferink, H.; Adema, G.J.; Boltje, T.J. Potent Metabolic Sialylation Inhibitors Based on C-5-Modified Fluorinated Sialic Acids. J. Med. Chem. 2019, 62, 1014–1021. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A Summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.Q.; Jiang, Z.Z.; Li, L.; Wu, Y.; Zheng, L. CD169 Identifies an Anti-Tumour Macrophage Subpopulation in Human Hepatocellular Carcinoma. J. Pathol. 2016, 239, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zeng, D.N.; Li, J.Z.; Hua, Q.M.; Huang, C.X.; Xu, J.; Wu, C.; Zheng, L.; Wen, W.P.; Wu, Y. Type I IFNs Repolarized a CD169+ Macrophage Population with Anti-Tumor Potentials in Hepatocellular Carcinoma. Mol. Ther. 2022, 30, 632–643. [Google Scholar] [CrossRef]
- Jing, W.; Guo, X.; Wang, G.; Bi, Y.; Han, L.; Zhu, Q.; Qiu, C.; Tanaka, M.; Zhao, Y. Breast Cancer Cells Promote CD169+ Macrophage-Associated Immunosuppression through JAK2-Mediated PD-L1 Upregulation on Macrophages. Int. Immunopharmacol. 2020, 78, 106012. [Google Scholar] [CrossRef] [PubMed]
- Lindau, D.; Gielen, P.; Kroesen, M.; Wesseling, P.; Adema, G.J. The Immunosuppressive Tumour Network: Myeloid-Derived Suppressor Cells, Regulatory T Cells and Natural Killer T Cells. Immunology 2013, 138, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Dusoswa, S.A.; Verhoeff, J.; Abels, E.; Méndez-Huergo, S.P.; Croci, D.O.; Kuijper, L.H.; de Miguel, E.; Wouters, V.M.C.J.; Best, M.G.; Rodriguez, E.; et al. Glioblastomas Exploit Truncated O-Linked Glycans for Local and Distant Immune Modulation via the Macrophage Galactose-Type Lectin. Proc. Natl. Acad. Sci. USA 2020, 117, 3693–3703. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Xia, Y.; Bettegowda, C.; Weller, M. Current State of Immunotherapy for Glioblastoma. Nat. Rev. Clin. Oncol. 2018, 15, 422–442. [Google Scholar] [CrossRef] [PubMed]
- Gielen, P.; Van Der Valk, P.; Wesseling, P. Immune Responses to Tumours in the CNS. In Neuroinflammation and CNS Disorders; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 363–384. [Google Scholar] [CrossRef]
- Maniecki, M.B.; Etzerodt, A.; Ulhøi, B.P.; Steiniche, T.; Borre, M.; Dyrskjøt, L.; Ørntoft, T.F.; Moestrup, S.K.; Møller, H.J. Tumor-Promoting Macrophages Induce the Expression of the Macrophage-Specific Receptor CD163 in Malignant Cells. Int. J. Cancer 2012, 131, 2320–2331. [Google Scholar] [CrossRef] [PubMed]
- Medrek, C.; Pontén, F.; Jirström, K.; Leandersson, K. The Presence of Tumor Associated Macrophages in Tumor Stroma as a Prognostic Marker for Breast Cancer Patients. BMC Cancer 2012, 12, 306. [Google Scholar] [CrossRef]
- Chen, T.; Chen, J.; Zhu, Y.; Li, Y.; Wang, Y.; Chen, H.; Wang, J.; Li, X.; Liu, Y.; Li, B.; et al. CD163, a Novel Therapeutic Target, Regulates the Proliferation and Stemness of Glioma Cells via Casein Kinase 2. Oncogene 2018, 38, 1183–1199. [Google Scholar] [CrossRef]
- Yang, F.; He, Z.; Duan, H.; Zhang, D.; Li, J.; Yang, H.; Dorsey, J.F.; Zou, W.; Ali Nabavizadeh, S.; Bagley, S.J.; et al. Synergistic Immunotherapy of Glioblastoma by Dual Targeting of IL-6 and CD40. Nat. Commun. 2021, 12, 3424. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cornelissen, L.A.M.; Santegoets, K.C.M.; Kers-Rebel, E.D.; Bossmann, S.A.J.F.H.; Ter Laan, M.; Granado, D.; Adema, G.J. Glioma-Associated Sialoglycans Drive the Immune Suppressive Phenotype and Function of Myeloid Cells. Pharmaceutics 2024, 16, 953. https://doi.org/10.3390/pharmaceutics16070953
Cornelissen LAM, Santegoets KCM, Kers-Rebel ED, Bossmann SAJFH, Ter Laan M, Granado D, Adema GJ. Glioma-Associated Sialoglycans Drive the Immune Suppressive Phenotype and Function of Myeloid Cells. Pharmaceutics. 2024; 16(7):953. https://doi.org/10.3390/pharmaceutics16070953
Chicago/Turabian StyleCornelissen, Lenneke A. M., Kim C. M. Santegoets, Esther D. Kers-Rebel, Sandra A. J. F. H. Bossmann, Mark Ter Laan, Daniel Granado, and Gosse J. Adema. 2024. "Glioma-Associated Sialoglycans Drive the Immune Suppressive Phenotype and Function of Myeloid Cells" Pharmaceutics 16, no. 7: 953. https://doi.org/10.3390/pharmaceutics16070953