
RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Physiological Responses
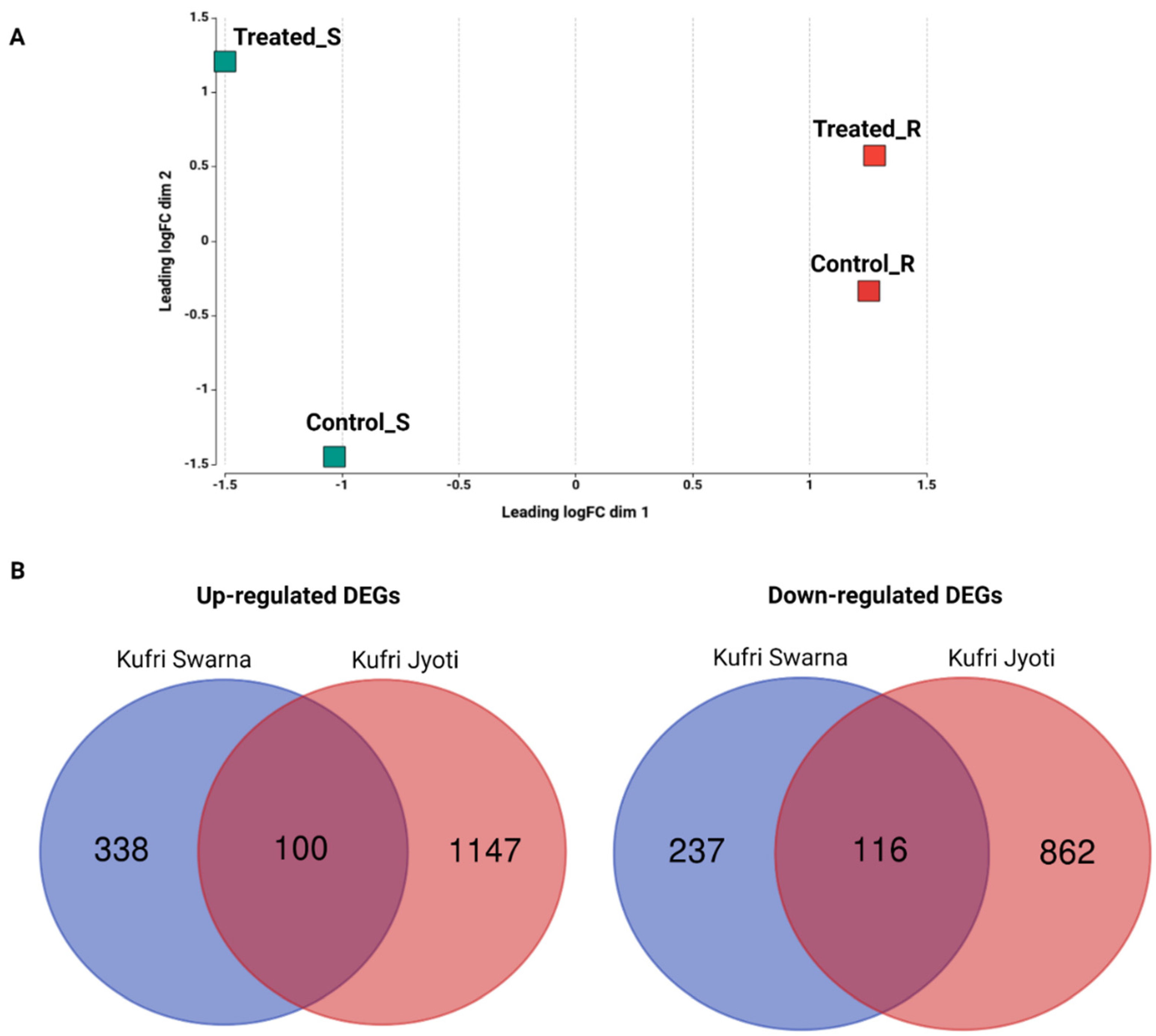
2.2. RNA-seq and Analysis
2.3. Differentially Expressed Genes
2.4. Pathway Enrichment Analysis
2.5. Biotic Stress Pathway
2.6. DEGs for TFs and Disease Resistance
2.7. Gene Ontology (GO) Enrichment Analysis of DEGs
2.8. Functional Network Analysis of DEGs
2.9. Validation of RNA-seq Results with qRT-PCR Analysis
3. Discussion
4. Materials and Methods
4.1. In Vitro Propagation of PCN-Resistant and -Susceptible Cultivars
4.2. PCN Hatching and Inoculation
4.3. RNA Isolation, Library Preparation, and RNA-seq
4.4. Preprocessing and Genome Mapping
4.5. Identification of Differentially Expressed Genes (DEGs)
4.6. qRT-PCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Zhang, X.; Na, R.; Yang, S.; Tian, Z.; Zhao, Y.; Zhao, J. StRac1 Plays an Important Role in Potato Resistance Against Phytophthora Infestans Via Regulating H2O2 Production. J. Plant Physiol. 2020, 253, 153249. [Google Scholar] [CrossRef] [PubMed]
- Sudha, R.; Mhatre, P.; Lekshmanan, D.; Venkatasalam, E.; Bairwa, A.; Bhardwaj, V.; Dalamu; Sharma, R. Phenotypic and Molecular Characterization of Potato Germplasm for Potato Cyst Nematode Resistance. Indian J. Genet. 2019, 79, 394–403. [Google Scholar] [CrossRef]
- Mishra, T.; Raigond, P.; Thakur, N.; Dutt, S.; Singh, B. Recent Ipdates on Healthy Phytoconstituents in Potato: A Nutritional Depository. Potato Res. 2020, 63, 323–343. [Google Scholar] [CrossRef]
- Munnaf, M.A.; Haesaert, G.; Van Meirvenne, M.; Mouazen, A.M. Map-Based Site-Specific Seeding of Consumption Potato Production Using High-Resolution Soil and Crop Data Fusion. Comput. Electron. Agric. 2020, 178, 105752. [Google Scholar] [CrossRef]
- Seenivasan, N. Status of Potato Cyst Nematodes Globodera spp. Infection on Potato at Kodaikanal Hill of Tamil Nadu, India and Yield Loss Estimation. J. Entomol. Zoo Stud. 2017, 5, 268–272. [Google Scholar]
- Mhatre, P.H.; Divya, K.; Venkatasalam, E.; Bairwa, A.; Sudha, R.; Berliner, J. Potato Cyst Nematode: A hidden Enemy of Potato Cultivation in Hills. Bhartiya Krishi Anusandhan Patrika 2019, 34, 50–53. [Google Scholar] [CrossRef]
- Bairwa, A.; Venkatasalam, E.; Jeevalatha, A.; Priyank, H.; Buckseth, T.; Jenifer, A.; Sharma, S.; Singh, R.; Chakrabarti, S. Morphological and Molecular Characterization of Potato Cyst Nematode Populations from the Nilgiris. Indian J. Agric. Sci. 2020, 90, 273–278. [Google Scholar]
- Gavrilenko, T.A.; Khiutti, A.V.; Klimenko, N.S.; Antonova, O.Y.; Fomina, N.A.; Afanasenko, O.S. Phenotypic and DNA Marker-Assisted Characterization of Russian Potato Cultivars for Resistance to Potato Cyst Nematodes. Agronomy 2021, 11, 2400. [Google Scholar] [CrossRef]
- Mburu, H.; Cortada, L.; Haukeland, S.; Ronno, W.; Nyongesa, M.; Kinyua, Z.; Bargul, J.L.; Coyne, D. Potato Cyst Nematodes: A New Threat to Potato Production in East Africa. Front. Plant Sci. 2020, 11, 670. [Google Scholar] [CrossRef]
- Varandas, R.; Egas, C.; Conceição, I.L. Potato Cyst Nematodes: New Solutions to an Old Problem. Crop. Prot. 2020, 137, 105303. [Google Scholar] [CrossRef]
- Bairwa, A.; Venkatasalam, E.; Sudha, R.; Umamaheswari, R.; Singh, B. Techniques for Characterization and Eradication of Potato Cyst Nematode: A Review. J. Parasit. Dis. 2017, 41, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Nowaczyk, K.; Dobosz, R.; Budziszewska, M.; Kamasa, J.; ObrĘpalska-StĘplowska, A. Analysis of Diversity of Golden Potato Cyst Nematode (Globodera rostochiensis) Populations from Poland Using Molecular Approaches. J. Phytopathol. 2011, 159, 759–766. [Google Scholar] [CrossRef]
- Mhatre, P.H.; Divya, K.; Venkatasalam, E.; Bairwa, A.; Sudha, R.; Saranya, C.; Guru-Pirasanna-Pandi, G.; Sharma, S. Evaluation of Trap Crop, Solanum Sisymbriifolium and Antagonistic Crops Against Potato Cyst Nematodes, Globodera spp. S. Afr. J. Bot. 2021, 138, 242–248. [Google Scholar] [CrossRef]
- Chandel, Y.; Bhadu, S.S.; Salalia, R.; Thakur, S.; Kumar, S.; Somvanshi, V.S.; Mukherjee, A.; Walia, R. Prevalence and Spread of Potato Cyst Nematodes, Globodera spp. in Northern Hilly Areas of India. Curr. Sci. 2020, 118, 1946. [Google Scholar] [CrossRef]
- López-Lima, D.; Desgarennes, D.; Lima-Rivera, D.; Carrión, G. Integrated Management of Globodera Rostochiensis: A Novel Biocontrol Agent, Crop Rotation and Fallow. J. Plant Dis. Prot. 2020, 127, 633–640. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Swapnil, P. Plant Growth-Promoting Rhizobacteria as a Green Alternative for Sustainable Agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Wang, N.; Wang, L.; Zhu, K.; Hou, S.; Chen, L.; Mi, D.; Gui, Y.; Qi, Y.; Jiang, C.; Guo, J.H. Plant Root Exudates Are Involved in Bacillus Cereus AR156 Mediated Biocontrol Against Ralstonia Solanacearum. Front. Microbiol. 2019, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Putker, V.; Goverse, A. Molecular and Cellular Mechanisms Involved in Host-Specific Resistance to Cyst Nematodes in Crops. Front. Plant Sci. 2021, 12, 363. [Google Scholar] [CrossRef]
- Santini, A.; Battisti, A. Complex Insect–Pathogen Interactions in Tree Pandemics. Front. Physiol. 2019, 10, 550. [Google Scholar] [CrossRef]
- Joshi, I.; Kohli, D.; Pal, A.; Chaudhury, A.; Sirohi, A.; Jain, P.K. Host Delivered-RNAi of Effector Genes for Imparting Resistance Against Root-Knot and Cyst Nematodes in Plants. Physiol. Mol. Plant Pathol. 2022, 118, 101802. [Google Scholar] [CrossRef]
- Thordal-Christensen, H. A Holistic View on Plant Effector-Triggered Immunity Presented as an Iceberg Model. Cell. Mol. Life Sci. 2020, 77, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, Y.; Fudal, I. Complex Interactions between Fungal Avirulence Genes and Their Corresponding Plant Resistance Genes and Consequences for Disease Resistance Management. Front. Plant Sci. 2017, 8, 1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, P.; Gleason, C. Plant-Parasitic Nematode Effectors—Insights Into Their Diversity and New Tools for Their Identification. Curr. Opin. Plant Biol. 2019, 50, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Lightbody, G.; Haberland, V.; Browne, F.; Taggart, L.; Zheng, H.; Parkes, E.; Blayney, J.K. Review of Applications of High-Throughput Sequencing in Personalized Medicine: Barriers and Facilitators of Future Progress in Research and Clinical Application. Brief. Bioinform. 2019, 20, 1795–1811. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Sun, M.A.; Yan, P. Pathway and Network Analysis of Differentially Expressed Genes in Transcriptomes. In Transcriptome Data Analysis; Springer: Berlin/Heidelberg, Germany, 2018; pp. 35–55. [Google Scholar]
- Shukla, N.; Yadav, R.; Kaur, P.; Rasmussen, S.; Goel, S.; Agarwal, M.; Jagannath, A.; Gupta, R.; Kumar, A. Transcriptome Analysis of Root-Knot Nematode (Meloidogyne Incognita)-Infected Tomato (Solanum Lycopersicum) Roots Reveals Complex Gene Expression Profiles and Metabolic Networks of Both Host and Nematode During Susceptible and Resistance Responses. Mol. Plant Pathol. 2018, 19, 615–633. [Google Scholar] [CrossRef] [Green Version]
- Cao, K.; Li, H.; Wang, Q.; Zhao, P.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wang, L. Comparative Transcriptome Analysis of Genes Involved in the Response of Resistant and Susceptible Peach Cultivars to Nematode Infection. Sci. Hortic. 2017, 215, 20–27. [Google Scholar] [CrossRef]
- Xing, X.; Li, X.; Zhang, M.; Wang, Y.; Liu, B.; Xi, Q.; Zhao, K.; Wu, Y.; Yang, T. Transcriptome Analysis of Resistant and Susceptible Tobacco (Nicotiana Tabacum) in Response to Root-Knot Nematode Meloidogyne Incognita Infection. Biochem. Biophys. Res. Commun. 2017, 482, 1114–1121. [Google Scholar] [CrossRef]
- Kumari, C.; Dutta, T.K.; Banakar, P.; Rao, U. Comparing the Defence-Related Gene Expression Changes Upon Root-Knot Nematode Attack in Susceptible Versus Resistant Cultivars of Rice. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aditya, J.; Lewis, J.; Shirley, N.J.; Tan, H.T.; Henderson, M.; Fincher, G.B.; Burton, R.A.; Mather, D.E.; Tucker, M.R. The Dynamics of Cereal Cyst Nematode Infection Differ between Susceptible and Resistant Barley Cultivars and Lead to Changes in (1,3;1,4)-β-Glucan Levels and HvCslF Gene Transcript Abundance. New Phytol. 2015, 207, 135–147. [Google Scholar] [CrossRef]
- Walter, A.J.; Willforss, J.; Lenman, M.; Alexandersson, E.; Andreasson, E. RNA Seq Analysis of Potato Cyst Nematode Interactions with Resistant and Susceptible Potato Roots. Eur. J. Plant Pathol. 2018, 152, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Indian Potato Varieties and Their Salient Features; ICAR-Central Potato Research Institute: Shimla, India, 2014. [Google Scholar]
- Sudha, R.; Venkatasalam, E.; Bairwa, A.; Bhardwaj, V.; Sharma, R. Identification of Potato Cyst Nematode Resistant Genotypes Using Molecular Markers. Sci. Hortic. 2016, 198, 21–26. [Google Scholar] [CrossRef]
- Pham, G.M.; Hamilton, J.P.; Wood, J.C.; Burke, J.T.; Zhao, H.; Vaillancourt, B.; Ou, S.; Jiang, J.; Buell, C.R. Construction of A Chromosome-Scale Long-Read Reference Genome Assembly for Potato. Gigascience 2020, 9, giaa100. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, S. MAPK Cascades in Plant Disease Resistance Signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional Roles of Plant Cuticle During Plant-Pathogen Interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Anjam, M.S.; Nawaz, M.A.; Lam, H.M.; Chung, G. Signal Transduction in Plant–Nematode Interactions. Int. J. Mol. Sci. 2018, 19, 1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Chittem, K.; Brueggeman, R.; Osorno, J.M.; Richards, J.; Nelson, B.D. Comparative Transcriptome Analysis of Resistant and Susceptible Common Bean Genotypes in Response to Soybean Cyst Nematode Infection. PLoS ONE 2016, 11, e0159338. [Google Scholar]
- Kumar, P.; Khanal, S.; Da Silva, M.; Singh, R.; Davis, R.F.; Nichols, R.L.; Chee, P.W. Transcriptome Analysis of a Nematode Resistant and Susceptible Upland Cotton Line at Two Critical Stages of Meloidogyne Incognita Infection and Development. PLoS ONE 2019, 14, e0221328. [Google Scholar] [CrossRef]
- Sato, K.; Uehara, T.; Holbein, J.; Sasaki-Sekimoto, Y.; Gan, P.; Bino, T.; Yamaguchi, K.; Ichihashi, Y.; Maki, N.; Shigenobu, S. Transcriptomic Analysis of Resistant and Susceptible Responses in a New Model Root-Knot Nematode Infection System Using Solanum Torvum and Meloidogyne Arenaria. Front. Plant Sci. 2021, 12, 680151. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant Hormone-Mediated Regulation of Stress Responses. BMC Plant Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ithal, N.; Recknor, J.; Nettleton, D.; Maier, T.; Baum, T.J.; Mitchum, M.G. Developmental Transcript Profiling of Cyst Nematode Feeding Cells in Soybean Roots. Mol. Plant Microbe Interact. 2007, 20, 510–525. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, T.; Tomitaka, Y.; Abe, H.; Tsuda, S.; Futai, K.; Mizukubo, T. Expression Profile of Jasmonic Acid-Induced Genes and the Induced Resistance Against The Root-Knot Nematode (Meloidogyne Incognita) in Tomato Plants (Solanum lycopersicum) after Foliar Treatment with Methyl Jasmonate. J. Plant Physiol. 2011, 168, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Maboreke, H.R.; Feldhahn, L.; Bönn, M.; Tarkka, M.T.; Buscot, F.; Herrmann, S.; Menzel, R.; Ruess, L. Transcriptome Analysis in Oak Uncovers A Strong Impact of Endogenous Rhythmic Growth on the Interaction with Plant-Parasitic Nematodes. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammerhofer, N.; Radakovic, Z.; Regis, J.M.; Dobrev, P.; Vankova, R.; Grundler, F.M.; Siddique, S.; Hofmann, J.; Wieczorek, K. Role of Stress-Related Hormones in Plant Defence During Early Infection of the Cyst Nematode Heterodera Schachtii in Arabidopsis. New Phytol. 2015, 207, 778–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhao, D.; Shuang, L.; Xiao, D.; Xuan, Y.; Duan, Y.; Chen, L.; Wang, Y.; Liu, X.; Fan, H. Transcriptome Analysis of Rice Roots in Response to Root-Knot Nematode Infection. Int. J. Mol. Sci. 2020, 21, 848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, K.K.; Xie, Q.G.; Mantelin, S.; Bishnoi, U.; Girke, T.; Navarre, D.A.; Kaloshian, I. Tomato Susceptibility to Root-Knot Nematodes Requires An Intact Jasmonic Acid Signaling Pathway. Mol. Plant Microbe Interact. 2008, 21, 1205–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, H.; Gheysen, G.; Denil, S.; Lindsey, K.; Topping, J.F.; Nahar, K.; Haegeman, A.; De Vos, W.H.; Trooskens, G.; Van Criekinge, W. Transcriptional Analysis Through RNA Sequencing of Giant Cells Induced by Meloidogyne Graminicola in Rice Roots. J. Exp. Bot. 2013, 64, 3885–3898. [Google Scholar] [CrossRef] [Green Version]
- Kyndt, T.; Fernandez, D.; Gheysen, G. Plant-Parasitic Nematode Infections in Rice: Molecular and Cellular Insights. Annu. Rev. Phytopathol. 2014, 52, 135–153. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone Crosstalk in Plant Disease and Defense: More Than Just Jasmonate-Salicylate Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Zhang, J.; Wen, Z.; Li, W.; Zhang, Y.; Zhang, L.; Dai, H.; Wang, D.; Xu, R. Genome-Wide Association Study for Soybean Cyst Nematode Resistance in Chinese Elite Soybean Cultivars. Mol. Breed. 2017, 37, 1–10. [Google Scholar] [CrossRef]
- Sidonskaya, E.; Schweighofer, A.; Shubchynskyy, V.; Kammerhofer, N.; Hofmann, J.; Wieczorek, K.; Meskiene, I. Plant Resistance Against the Parasitic Nematode Heterodera Schachtii is Mediated by MPK3 and MPK6 Kinases, Which are Controlled by the MAPK Phosphatase AP2C1 in Arabidopsis. J. Exp. Bot. 2016, 67, 107–118. [Google Scholar] [CrossRef] [Green Version]
- McNeece, B.T.; Sharma, K.; Lawrence, G.W.; Lawrence, K.S.; Klink, V.P. The Mitogen Activated Protein Kinase (MAPK) Gene Family Functions as a Cohort During the Glycine Max Defense Response to Heterodera glycines. Plant Physiol. Biochem. 2019, 137, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, Q. Poor Expression of MicroRNA-135b Results in the Inhibition of Cisplatin Resistance and Proliferation and Induces the Apoptosis of Gastric Cancer Cells Through MST1-Mediated MAPK Signaling Pathway. FASEB J. 2019, 33, 3420–3436. [Google Scholar] [CrossRef] [PubMed]
- Turra, D.; Bellin, D.; Lorito, M.; Gebhardt, C. Genotype-Dependent Expression of Specific Members of Potato Protease Inhibitor Gene Families in Different Tissues and in Response to Wounding and Nematode Infection. J. Plant Physiol. 2009, 166, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Zhu, X.; Wang, Y.; Chen, L.; Duan, Y. Transcriptomic and Metabolomic Analyses Reveal That Bacteria Promote Plant Defense During Infection of Soybean Cyst Nematode in Soybean. BMC Plant Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urwin, P.E.; Atkinson, H.J.; Waller, D.A.; McPherson, M.J. Engineered Oryzacystatin-I Expressed in Transgenic Hairy Roots Confers Resistance to Globodera pallida. Plant J. 1995, 8, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Samac, D.A.; Smigocki, A.C. Expression of Oryzacystatin I and II in Alfalfa Increases Resistance to the Root-Lesion Nematode. Phytopathology 2003, 93, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Torres, J.L.; Wilbers, R.H.; Gawronski, P.; Boshoven, J.C.; Finkers-Tomczak, A.; Cordewener, J.H.; America, A.H.; Overmars, H.A.; Van‘t Klooster, J.W.; Baranowski, L. Dual Disease Resistance Mediated by the Immune Receptor Cf-2 in Tomato Requires a Common Virulence Target of a Fungus and a Nematode. Proc. Natl. Acad. Sci. USA 2012, 109, 10119–10124. [Google Scholar] [CrossRef] [Green Version]
- Jammes, F.; Lecomte, P.; de Almeida-Engler, J.; Bitton, F.; Martin-Magniette, M.L.; Renou, J.P.; Abad, P.; Favery, B. Genome-Wide Expression Profiling of the Host Response to Root-Knot Nematode Infection in Arabidopsis. Plant J. 2005, 44, 447–458. [Google Scholar] [CrossRef]
- Tirumalaraju, S.V.; Jain, M.; Gallo, M. Differential Gene Expression in Roots of Nematode-Resistant and-Susceptible Peanut (Arachis hypogaea) Cultivars in Response to Early Stages of Peanut Root-Knot Nematode (Meloidogyne arenaria) Parasitization. J. Plant Physiol. 2011, 168, 481–492. [Google Scholar] [CrossRef]
- Swiecicka, M.; Filipecki, M.; Lont, D.; Van Vliet, J.; Qin, L.; Goverse, A.; Bakker, J.; Helder, J. Dynamics in the Tomato Root Transcriptome on Infection with the Potato Cyst Nematode Globodera rostochiensis. Mol. Plant Pathol. 2009, 10, 487–500. [Google Scholar] [CrossRef]
- Hewezi, T.; Piya, S.; Richard, G.; Rice, J.H. Spatial and Temporal Expression Patterns of Auxin Response Transcription Factors in the Syncytium Induced by the Beet Cyst Nematode Heterodera schachtii in Arabidopsis. Mol. Plant Pathol. 2014, 15, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Beneventi, M.A.; da Silva, O.B.; de Sá, M.E.L.; Firmino, A.A.P.; de Amorim, R.M.S.; Albuquerque, É.V.S.; da Silva, M.C.M.; da Silva, J.P.; Campos, M.d.A.; Lopes, M.J.C. Transcription Profile of Soybean-Root-Knot Nematode Interaction Reveals a Key Role of Phythormones in the Resistance Reaction. BMC Genom. 2013, 14, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.A.; Azeem, F.; Li, H.; Bohlmann, H. Smart Parasitic Nematodes Use Multifaceted Strategies to Parasitize Plants. Front. Plant Sci. 2017, 8, 1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goellner, M.; Wang, X.; Davis, E.L. Endo-β-1, 4-Glucanase Expression in Compatible Plant–Nematode Interactions. Plant Cell 2001, 13, 2241–2255. [Google Scholar]
- Puthoff, D.P.; Nettleton, D.; Rodermel, S.R.; Baum, T.J. Arabidopsis Gene Expression Changes During Cyst Nematode Parasitism Revealed by Statistical Analyses of Microarray Expression Profiles. Plant J. 2003, 33, 911–921. [Google Scholar] [CrossRef]
- Szakasits, D.; Heinen, P.; Wieczorek, K.; Hofmann, J.; Wagner, F.; Kreil, D.P.; Sykacek, P.; Grundler, F.M.; Bohlmann, H. The Transcriptome of Syncytia Induced by the Cyst Nematode Heterodera schachtii in Arabidopsis Roots. Plant J. 2009, 57, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, K. Cell Wall Alterations in Nematode-Infected Roots. In Advances in Botanical Research; Elsevier: Madrid, Spain, 2015; Volume 73, pp. 61–90. [Google Scholar]
- Ithal, N.; Recknor, J.; Nettleton, D.; Hearne, L.; Maier, T.; Baum, T.J.; Mitchum, M.G. Parallel Genome-Wide Expression Profiling of Host and Pathogen During Soybean Cyst Nematode Infection of Soybean. Mol. Plant-Microbe Interact. 2007, 20, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Goverse, A.; Smant, G. The Activation and Suppression of Plant Innate Immunity by Parasitic Nematodes. Annu. Rev. Phytopathol. 2014, 52, 243–265. [Google Scholar] [CrossRef]
- Edwards, R.; Mizen, T.; Cook, R. Isoflavonoid Conjugate Accumulation in the Roots of Lucerne (Medicago sativa) Seedlings Following Infection by the Stem Nematode (Ditylenchus dipsaci). Nematologica 1995, 41, 51–66. [Google Scholar] [CrossRef]
- Chin, S.; Behm, C.A.; Mathesius, U. Functions of Flavonoids in Plant–Nematode Interactions. Plants 2018, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Vlachopoulos, E.; Smith, L. Flavonoids in Potato Cyst Nematodes. Fundam. Appl. Nematol. 1993, 16, 103–106. [Google Scholar]
- Hung, C.; Rohde, R. Phenol Accumulation Related to Resistance in Tomato to Infection by Root-Knot and Lesion Nematodes. J. Nematol. 1973, 5, 253. [Google Scholar] [PubMed]
- Postnikova, O.A.; Hult, M.; Shao, J.; Skantar, A.; Nemchinov, L.G. Transcriptome Analysis of Resistant and Susceptible Alfalfa Cultivars Infected with Root-Knot Nematode Meloidogyne incognita. PLoS ONE 2015, 10, e0118269. [Google Scholar] [CrossRef] [PubMed]
- Kandoth, P.K.; Ithal, N.; Recknor, J.; Maier, T.; Nettleton, D.; Baum, T.J.; Mitchum, M.G. The Soybean Rhg1 Locus for Resistance to the Soybean Cyst Nematode Heterodera glycines Regulates the Expression of a Large Number of Stress-and Defense-Related Genes in Degenerating Feeding Cells. Plant Physiol. 2011, 155, 1960–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, T.; Sugiyama, S.; Matsuura, H.; Arie, T.; Masuta, C. Resistant and susceptible Responses in Tomato to Cyst Nematode are Differentially Regulated by Salicylic Acid. Plant Cell Physiol. 2010, 51, 1524–1536. [Google Scholar] [CrossRef]
- Simonetti, E.; Alba, E.; Montes, M.J.; Delibes, A.; López-Braña, I. Analysis of Ascorbate Peroxidase Genes Expressed in Resistant and Susceptible Wheat Lines Infected by the Cereal Cyst Nematode, Heterodera avenae. Plant Cell Rep. 2010, 29, 1169–1178. [Google Scholar] [CrossRef]
- Chen, S.; Chronis, D.; Wang, X. The novel GrCEP12 Peptide from the Plant-Parasitic Nematode Globodera rostochiensis Suppresses Flg22-Mediated PTI. Plant Signal. Behav. 2013, 8, e25359. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.; Robertson, W.M.; Sobczak, M.; Helder, J.; Tetaud, E.; Ariyanayagam, M.R.; Ferguson, M.A.; Fairlamb, A.; Jones, J.T. Cloning, Expression and Functional Characterization of a Peroxiredoxin from the Potato Cyst Nematode Globodera Rostochiensis. Mol. Biochem. Parasitol. 2000, 111, 41–49. [Google Scholar] [CrossRef]
- Lee, C.; Chronis, D.; Kenning, C.; Peret, B.; Hewezi, T.; Davis, E.L.; Baum, T.J.; Hussey, R.; Bennett, M.; Mitchum, M.G. The Novel Cyst Nematode Effector Protein 19C07 Interacts with the Arabidopsis Auxin Influx Transporter LAX3 to Control Feeding Site Development. Plant Physiol. 2011, 155, 866–880. [Google Scholar] [CrossRef] [Green Version]
- Doyle, E.A.; Lambert, K.N. Meloidogyne Javanica Chorismate Mutase 1 Alters Plant Cell Development. Mol. Plant-Microbe Interact. 2003, 16, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Clarke, A.; Perry, R. Hatching of Cyst-Nematodes. Nematologica 1977, 23, 350–368. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—APython Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Tarazona, S.; Furió-Tarí, P.; Turrà, D.; Pietro, A.D.; Nueda, M.J.; Ferrer, A.; Conesa, A. Data Quality Aware Analysis of Differential Expression in RNA-Seq with NOISeq R/Bioc Package. Nucleic Acids Res. 2015, 43, e140. [Google Scholar] [CrossRef] [Green Version]
- Osuna-Cruz, C.M.; Paytuvi-Gallart, A.; Di Donato, A.; Sundesha, V.; Andolfo, G.; Aiese Cigliano, R.; Sanseverino, W.; Ercolano, M.R. PRGdb 3.0: A Comprehensive Platform for Prediction and Analysis of Plant Disease Resistance Genes. Nucleic Acids Res. 2018, 46, D1197–D1201. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particulars | Control_S | Treated_S | Control_R | Treated_R |
|---|---|---|---|---|
| Total number of raw reads | 20,099,283 | 20,535,765 | 21,367,409 | 24,128,453 |
| Total number of valid paired-end reads | 19,116,371 | 19,544,076 | 19,976,274 | 23,014,160 |
| Read length | 75 | 75 | 75 | 75 |
| GC content (%) | 42 | 42 | 42 | 42 |
| Q30 (%) | 95.1 | 95.2 | 93.5 | 95.4 |
| % of mapped reads | 97.3 | 96.4 | 86.1 | 88.9 |
| % of unmapped reads | 2.7 | 3.6 | 13.92 | 11.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrasekar, S.; Natarajan, P.; Mhatre, P.H.; Mahajan, M.; Nivitha, S.; Palanisamy, V.E.; Reddy, U.K.; Sundararaj, P. RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars. Plants 2022, 11, 1008. https://doi.org/10.3390/plants11081008
Chandrasekar S, Natarajan P, Mhatre PH, Mahajan M, Nivitha S, Palanisamy VE, Reddy UK, Sundararaj P. RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars. Plants. 2022; 11(8):1008. https://doi.org/10.3390/plants11081008
Chicago/Turabian StyleChandrasekar, Saranya, Purushothaman Natarajan, Priyank Hanuman Mhatre, Mahesh Mahajan, Sundararaj Nivitha, Venkatasalam E. Palanisamy, Umesh K. Reddy, and Palanisamy Sundararaj. 2022. "RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars" Plants 11, no. 8: 1008. https://doi.org/10.3390/plants11081008
APA StyleChandrasekar, S., Natarajan, P., Mhatre, P. H., Mahajan, M., Nivitha, S., Palanisamy, V. E., Reddy, U. K., & Sundararaj, P. (2022). RNA-Seq of Cyst Nematode Infestation of Potato (Solanum tuberosum L.): A Comparative Transcriptome Analysis of Resistant and Susceptible Cultivars. Plants, 11(8), 1008. https://doi.org/10.3390/plants11081008