
Thaumatin-like Proteins in Legumes: Functions and Potential Applications—A Review




Abstract
1. Introduction
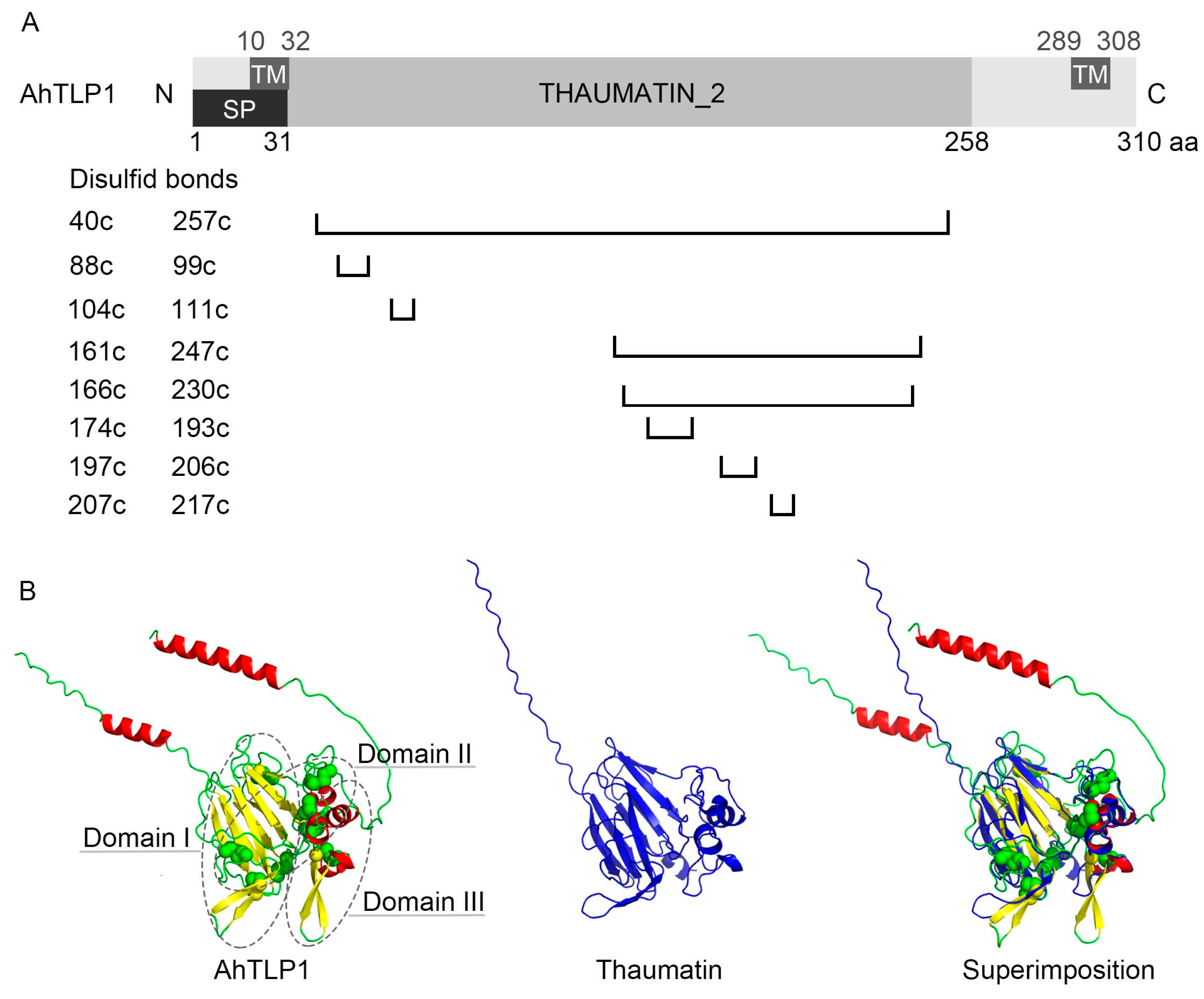
2. Classification and Characteristics of TLPs
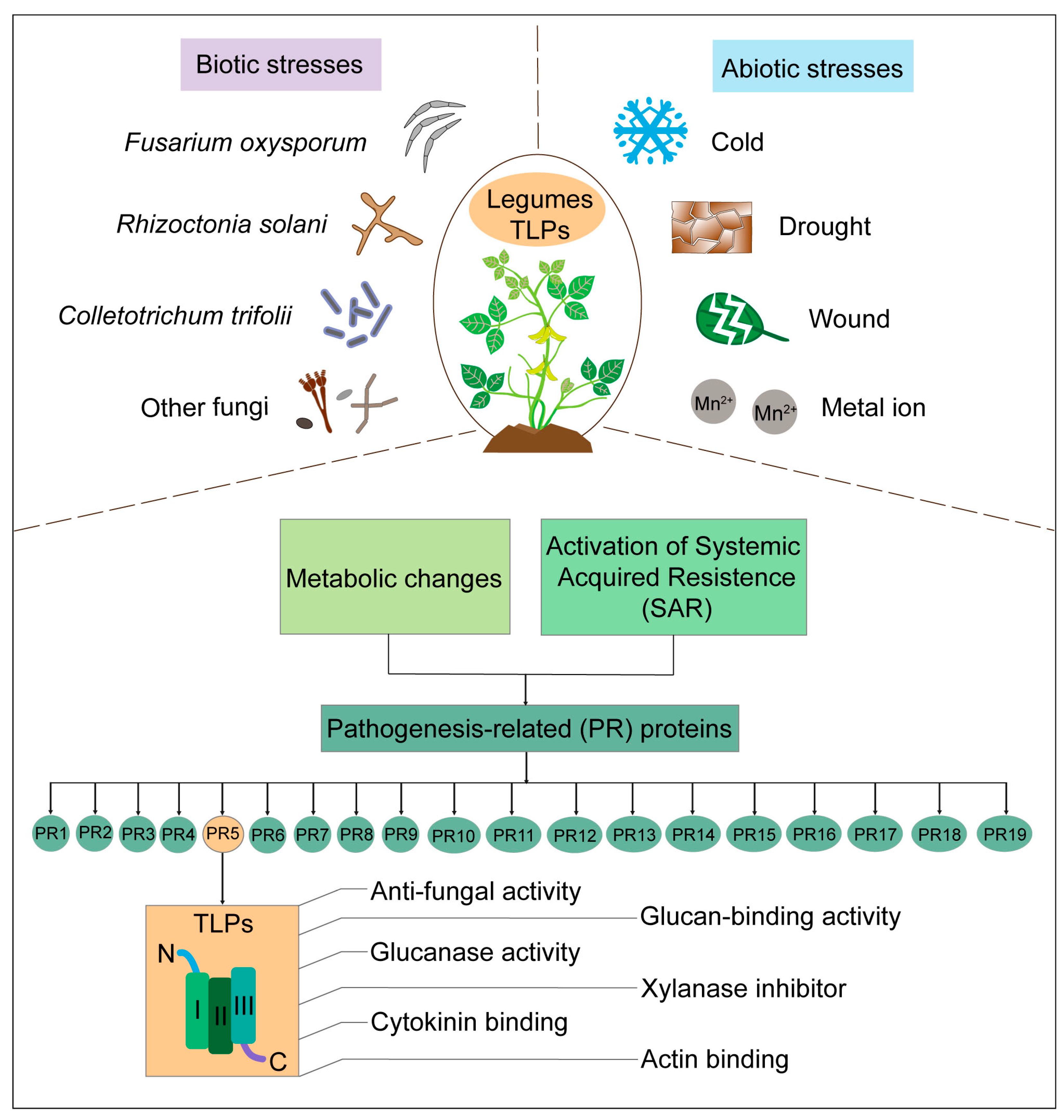
3. Biological Functions of TLPs in Legumes
3.1. Legume TLPs in Response to Biotic Stresses
3.2. Legume TLPs in Response to Abiotic Stresses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sources | Function/Biological Role | References |
|---|---|---|---|
| TLP | Phaseolus vulgaris | Antifungal activity | [63] |
| CdTLP | Cassia didymobotrya | Antifungal activity | [59] |
| AdTLP | Arachis diogoi | Antifungal activity | [58] |
| MtTLP | Medicago truncatula | Antifungal activity | [56,64] |
| TLP | Cicer arietinum | Phytoalexin response | [65,66] |
| VfTLP4-3, VfTLP5 | Vicia faba | Drought response | [36] |
| StTLP1b | Senna tora | Wound response | [73] |
| AHCSP33 | Arachis hypogaea | Cold response | [32] |
| AnTLPs | Ammopiptanthus nanus | Cold response | [74] |
| GmOLPa | Glycine max | Salt response | [33,34] |
| GmOLPb | Glycine max | Salt response | [33,34] |
| TLP | Vigna unguiculata | Manganese toxicity response | [75] |
| TLP1b | Arachis hypogaea | Immune response | [72] |
| GmOLPa-like and P21-like osmotins | Glycine max | Drought response | [31] |
3.3. The Role of TLPs in Phytohormone Responses
3.4. The Role of TLPs in Leguminous Nodulation and Symbiosis
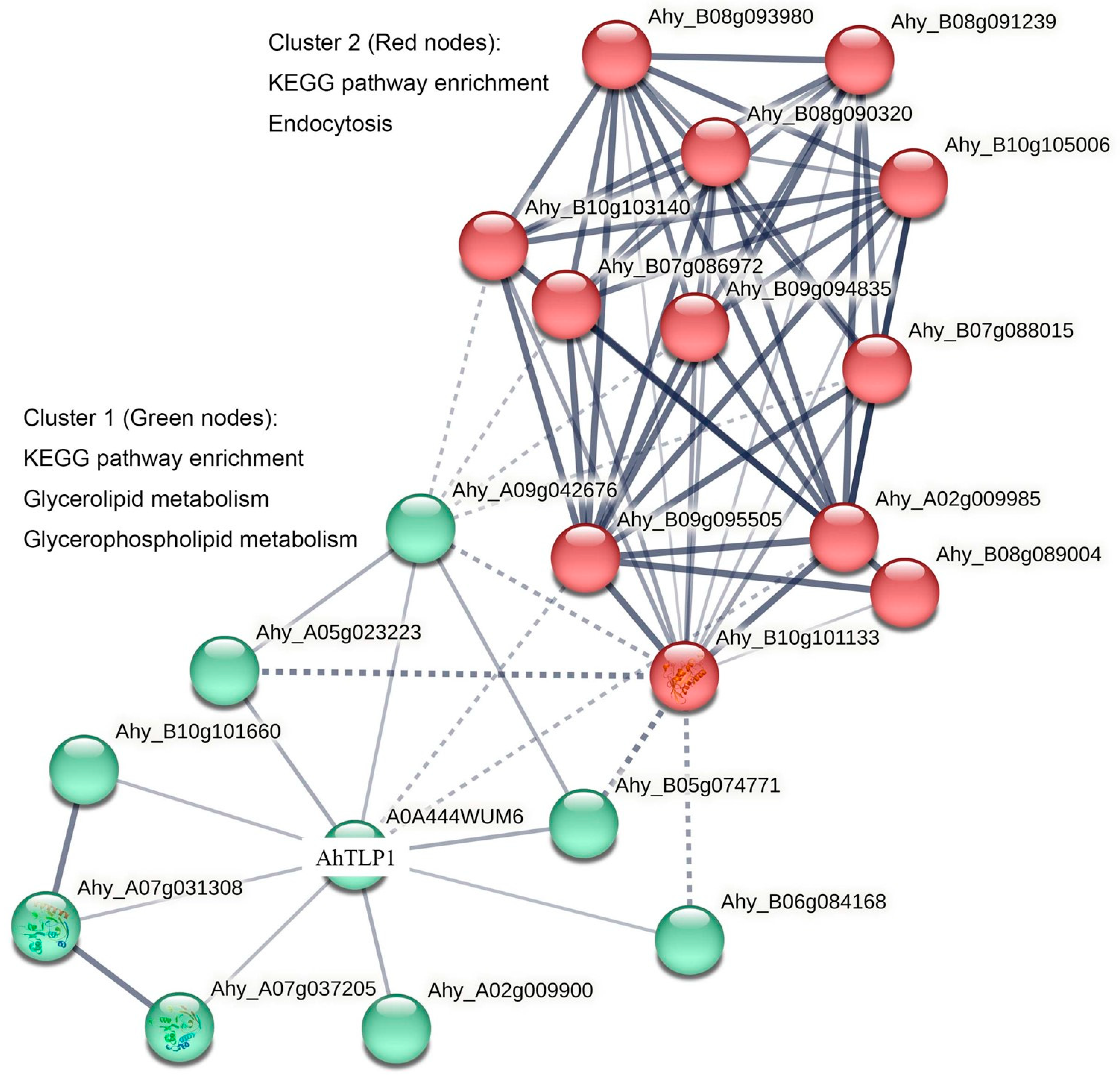
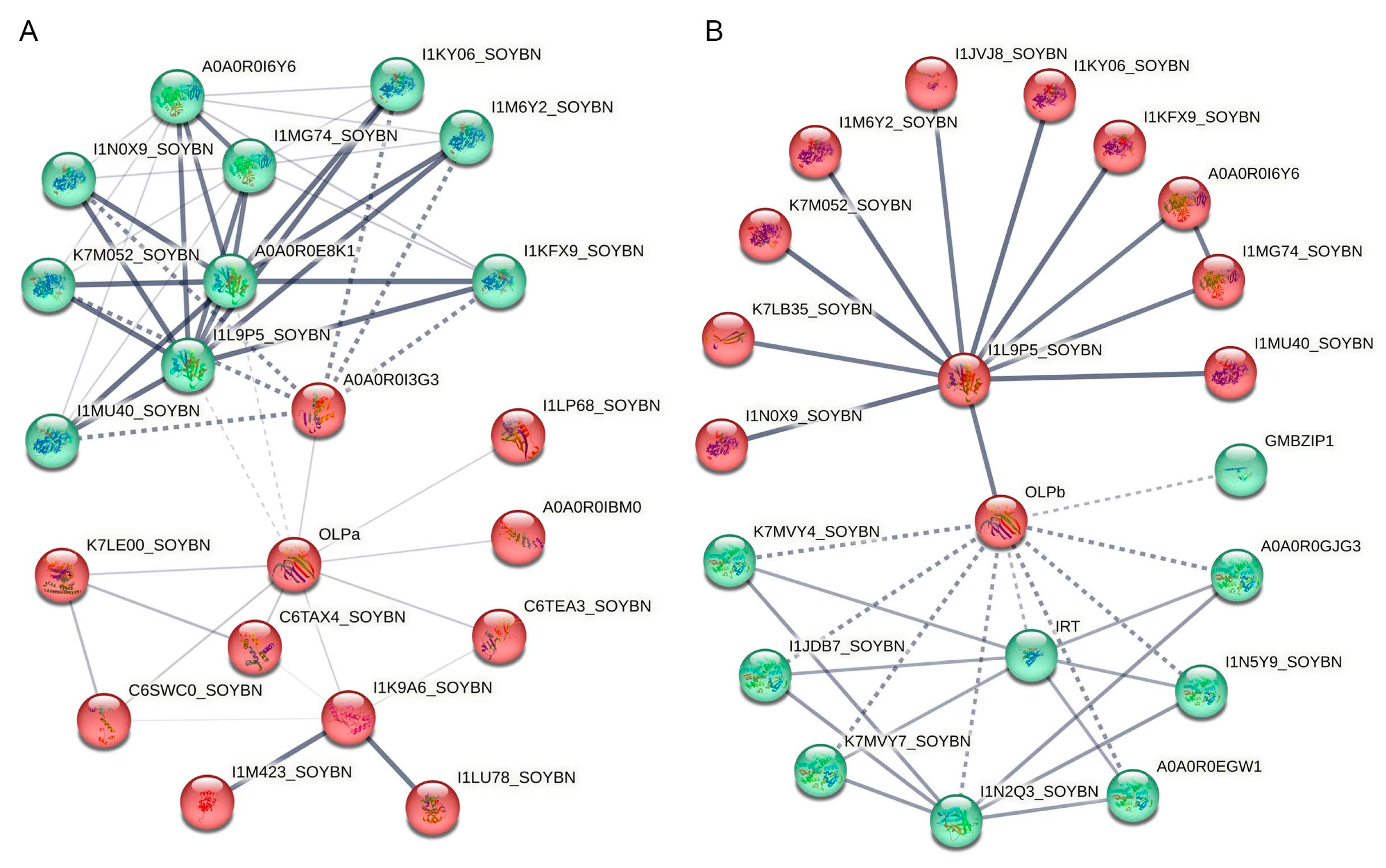
3.5. Prediction of Protein–Protein Interactions in Legume TLPs
4. Potential Biotechnological Applications for Legume TLPs
5. Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dos Santos, C.; Franco, O.L. Pathogenesis-Related Proteins (PRs) with Enzyme Activity Activating Plant Defense Responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Kou, Y.; Naqvi, N.I. Surface sensing and signaling networks in plant pathogenic fungi. Semin. Cell Dev. Biol. 2016, 57, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, B.; Xu, J.; Li, Z.; Jiang, C.; Zhou, Y.; Yang, Z.; Deng, M.; Lv, J.; Zhao, K. Tomato-Thaumatin-like Protein Genes Solyc08g080660 and Solyc08g080670 Confer Resistance to Five Soil-Borne Diseases by Enhancing beta-1,3-Glucanase Activity. Genes 2023, 14, 1622. [Google Scholar] [CrossRef]
- Liu, J.; Han, D.; Shi, Y. Gene Cloning, Expression, and Antifungal Activities of Permatin from Naked Oat (Avena nuda). Probiotics Antimicrob. Proteins 2019, 11, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, R.; Musidlak, O.; Barylski, J.; Nowicki, G.; Baldysz, S.; Czerwoniec, A.; Gozdzicka-Jozefiak, A. Characterization and expression of a novel thaumatin-like protein (CcTLP1) from papaveraceous plant Corydalis cava. Int. J. Biol. Macromol. 2021, 189, 678–689. [Google Scholar] [CrossRef]
- Zhou, H.; Xie, Y.; Jiang, Y.; Nadeem, H.; Wang, Y.; Yang, N.; Zhu, H.; Tang, C. GhTLP1, a thaumatin-like protein 1, improves Verticillium wilt resistance in cotton via JA, ABA and MAPK signaling pathway-plant pathways. Int. J. Biol. Macromol. 2023, 253, 127388. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, W.; Sang, X.; Wang, T.; Gong, H.; Zhao, Y.; Zhao, P.; Wang, H. Genome-Wide Identification of the Thaumatin-like Protein Family Genes in Gossypium barbadense and Analysis of Their Responses to Verticillium dahliae Infection. Plants 2021, 10, 2647. [Google Scholar] [CrossRef]
- Zhu, H.; Deng, M.; Yang, Z.; Mao, L.; Jiang, S.; Yue, Y.; Zhao, K. Two Tomato (Solanum lycopersicum) Thaumatin-Like Protein Genes Confer Enhanced Resistance to Late Blight (Phytophthora infestans). Phytopathology 2021, 111, 1790–1799. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, H.; He, S.; Zhang, P.; Ma, X. Genome-Wide Identification and Characterization of the TLP Gene Family in Phyllostachys edulis and Association with Witches’ Broom Disease Resistance in Bamboo. Int. J. Mol. Sci. 2023, 24, 10257. [Google Scholar] [CrossRef]
- Wang, T.; Li, X.; Zhang, C.; Xu, J. Transcriptome analysis of Ganoderma lingzhi (Agaricomycetes) response to Trichoderma hengshanicum infection. Front. Microbiol. 2023, 14, 1131599. [Google Scholar] [CrossRef] [PubMed]
- Gul, S.; Hussain, A.; Ali, Q.; Alam, I.; Alshegaihi, R.M.; Meng, Q.; Zaman, W.; Manghwar, H.; Munis, M.F.H. Hydropriming and Osmotic Priming Induce Resistance against Aspergillus niger in Wheat (Triticum aestivum L.) by Activating beta-1,3-glucanase, Chitinase, and Thaumatin-like Protein Genes. Life 2022, 12, 2061. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tang, C.; Deng, L.; Cai, G.; Liu, X.; Liu, B.; Han, Q.; Buchenauer, H.; Wei, G.; Han, D.; et al. Characterization of a pathogenesis-related thaumatin-like protein gene TaPR5 from wheat induced by stripe rust fungus. Physiol. Plant. 2010, 139, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xu, Y.; Wang, K.; Guo, W.; Gu, Y.; Lyu, S.; Huang, J.; Lin, H.; Huang, C.; Xu, Z.; et al. Genome-Wide Identification of TLP Gene Family and Their Roles in Carya cathayensis Sarg in Response to Botryosphaeria dothidea. Front. Plant Sci. 2022, 13, 849043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Miao, L.; Yang, X.; Jiang, G. Genome-wide characterization and expression of the TLP gene family associated with Colletotrichum gloeosporioides inoculation in Fragaria x ananassa. PeerJ 2022, 10, e12979. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, O.K.; Kochieva, E.Z.; Shchennikova, A.V.; Filyushin, M.A. Thaumatin-like Protein (TLP) Genes in Garlic (Allium sativum L.): Genome-Wide Identification, Characterization, and Expression in Response to Fusarium proliferatum Infection. Plants 2022, 11, 748. [Google Scholar] [CrossRef] [PubMed]
- Aloi, F.; Zamora-Ballesteros, C.; Martin-Garcia, J.; Diez, J.J.; Cacciola, S.O. Co-Infections by Fusarium circinatum and Phytophthora spp. on Pinus radiata: Complex Phenotypic and Molecular Interactions. Plants 2021, 10, 1976. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Fan, X.; Wang, G.; Xu, R.; Yan, L.; Zhou, Y.; Gong, Y.; Xiao, Y.; Bian, Y. Enhanced Expression of Thaumatin-like Protein Gene (LeTLP1) Endows Resistance to Trichoderma atroviride in Lentinula edodes. Life 2021, 11, 863. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, L.; Mi, X.; Zhao, S.; An, Y.; Xia, X.; Guo, R.; Wei, C. Multi-omics analysis to visualize the dynamic roles of defense genes in the response of tea plants to gray blight. Plant J. 2021, 106, 862–875. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, J.; Zhou, X.; Luan, Y.; Luan, F. Genome-wide identification, characterization and expression analysis of the TLP gene family in melon (Cucumis melo L.). Genomics 2020, 112, 2499–2509. [Google Scholar] [CrossRef]
- Jiao, W.; Li, X.; Zhao, H.; Cao, J.; Jiang, W. Antifungal Activity of an Abundant Thaumatin-Like Protein from Banana against Penicillium expansum, and Its Possible Mechanisms of Action. Molecules 2018, 23, 1442. [Google Scholar] [CrossRef] [PubMed]
- Odeny Ojola, P.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of rice thaumatin-like protein (Ostlp) gene in transgenic cassava results in enhanced tolerance to Colletotrichum gloeosporioides f. sp. manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Wu, J.; Zhang, Y.; Aguero, C.B.; Li, X.; Liu, S.; Wang, C.; Walker, M.A.; Lu, J. Overexpression of a thaumatin-like protein gene from Vitis amurensis improves downy mildew resistance in Vitis vinifera grapevine. Protoplasma 2017, 254, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Cao, Z.M.; Yu, Z.D.; Yu, D. Cloning and characterization of defense-related genes from Populus szechuanica infected with rust fungus Melampsora larici-populina. Genet. Mol. Res. 2016, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.; Pal, A.K.; Gulati, A.; Kumar, S.; Singh, A.K.; Ahuja, P.S. Overexpression of Camellia sinensis thaumatin-like protein, CsTLP in potato confers enhanced resistance to Macrophomina phaseolina and Phytophthora infestans infection. Mol. Biotechnol. 2013, 54, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Mani, T.; Sivakumar, K.C.; Manjula, S. Expression and functional analysis of two osmotin (PR5) isoforms with differential antifungal activity from Piper colubrinum: Prediction of structure-function relationship by bioinformatics approach. Mol. Biotechnol. 2012, 52, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, F.; Zhang, X.; Zhang, Y.; Hou, Y.; Zhang, S.; Wu, Z. Purification and characterization of a CkTLP protein from Cynanchum komarovii seeds that confers antifungal activity. PLoS ONE 2011, 6, e16930. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Isolation of an antifungal thaumatin-like protein from kiwi fruits. Phytochemistry 2002, 61, 1–6. [Google Scholar] [CrossRef]
- Lam, S.K.; Ng, T.B. First simultaneous isolation of a ribosome inactivating protein and an antifungal protein from a mushroom (Lyophyllum shimeji) together with evidence for synergism of their antifungal effects. Arch. Biochem. Biophys. 2001, 393, 271–280. [Google Scholar] [CrossRef]
- Faillace, G.R.; Caruso, P.B.; Timmers, L.; Favero, D.; Guzman, F.L.; Rechenmacher, C.; de Oliveira-Busatto, L.A.; de Souza, O.N.; Bredemeier, C.; Bodanese-Zanettini, M.H. Molecular Characterisation of Soybean Osmotins and Their Involvement in Drought Stress Response. Front. Genet. 2021, 12, 632685. [Google Scholar] [CrossRef]
- Dave, R.S.; Mitra, R.K. A low temperature induced apoplastic protein isolated from Arachis hypogaea. Phytochemistry 1998, 49, 2207–2213. [Google Scholar] [CrossRef] [PubMed]
- Tachi, H.; Fukuda-Yamada, K.; Kojima, T.; Shiraiwa, M.; Takahara, H. Molecular characterization of a novel soybean gene encoding a neutral PR-5 protein induced by high-salt stress. Plant Physiol. Biochem. 2009, 47, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Tachi, H.; Kojima, T.; Shiraiwa, M.; Takahara, H. Molecular cloning and characterization of a novel salt-inducible gene encoding an acidic isoform of PR-5 protein in soybean (Glycine max [L.] Merr.). Plant Physiol. Biochem. 2006, 44, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kirti, P.B. The genus Arachis: An excellent resource for studies on differential gene expression for stress tolerance. Front. Plant Sci. 2023, 14, 1275854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, X.; Zhang, J.; Huang, L.; Shi, Z.; Tian, Z.; Sha, A.; Lu, G. Thaumatin-like protein family genes VfTLP4-3 and VfTLP5 are critical for faba bean’s response to drought stress at the seedling stage. Plant Physiol. Biochem. 2023, 206, 108243. [Google Scholar] [CrossRef] [PubMed]
- Bohra, A.; Tiwari, A.; Kaur, P.; Ganie, S.A.; Raza, A.; Roorkiwal, M.; Mir, R.R.; Fernie, A.R.; Smykal, P.; Varshney, R.K. The Key to the Future Lies in the Past: Insights from Grain Legume Domestication and Improvement Should Inform Future Breeding Strategies. Plant Cell Physiol. 2022, 63, 1554–1572. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Sun, L.; Jiang, S.; Ren, H.; Sun, R.; Wei, Z.; Hong, H.; Luan, X.; Wang, J.; Wang, X.; et al. Soybean genetic resources contributing to sustainable protein production. Theor. Appl. Genet. 2022, 135, 4095–4121. [Google Scholar] [CrossRef] [PubMed]
- Shanthakumar, P.; Klepacka, J.; Bains, A.; Chawla, P.; Dhull, S.B.; Najda, A. The Current Situation of Pea Protein and Its Application in the Food Industry. Molecules 2022, 27, 5354. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Peanut (Arachis hypogaea L.): A Prospective Legume Crop to Offer Multiple Health Benefits Under Changing Climate. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1325–1338. [Google Scholar] [CrossRef]
- Mondal, S.; Badigannavar, A.M. Mapping of a dominant rust resistance gene revealed two R genes around the major Rust_QTL in cultivated peanut (Arachis hypogaea L.). Theor. Appl. Genet. 2018, 131, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, T.; Dai, X.; Yang, D.; Wu, Y.; Chen, H.; Zheng, Y.; Zhi, Q.; Wan, X.; Tan, X. Comparative transcriptome analysis revealed molecular mechanisms of peanut leaves responding to Ralstonia solanacearum and its type III secretion system mutant. Front. Microbiol. 2022, 13, 998817. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Smith, J.R. Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust. Plants 2023, 12, 1797. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, M.; Wang, L.; Feng, H.; He, X.; Chang, S.; Wang, D.; Wang, L.; Yang, J.; An, G.; et al. Whole-plant microbiome profiling reveals a novel geminivirus associated with soybean stay-green disease. Plant Biotechnol. J. 2022, 20, 2159–2173. [Google Scholar] [CrossRef] [PubMed]
- Lukanda, M.M.; Dramadri, I.O.; Adjei, E.A.; Badji, A.; Arusei, P.; Gitonga, H.W.; Wasswa, P.; Edema, R.; Ochwo-Ssemakula, M.; Tukamuhabwa, P.; et al. Genome-Wide Association Analysis for Resistance to Coniothyrium glycines Causing Red Leaf Blotch Disease in Soybean. Genes 2023, 14, 1271. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Liao, J.; He, Z.; Khurshid, M.; Wang, C.; Zhang, Z.; Mao, J.; Xia, Y. Effects of Peanut Rust Disease (Puccinia arachidis Speg.) on Agricultural Production: Current Control Strategies and Progress in Breeding for Resistance. Genes 2024, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, M.; Kemerait, R., Jr.; Bertioli, D.; Leal-Bertioli, S. Strong Resistance to Early and Late Leaf Spot in Peanut-Compatible Wild-Derived Induced Allotetraploids. Plant Dis. 2023, 107, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yang, F.; Hu, C.; Yang, X.; Zheng, A.; Wang, Y.; Tang, Y.; He, Y.; Lv, M. Production status and research advancement on root rot disease of faba bean (Vicia faba L.) in China. Front. Plant Sci. 2023, 14, 1165658. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Rastogi, A.; Yadav, S. Effects of Heat stress and molecular mitigation approaches in orphan legume, Chickpea. Mol. Biol. Rep. 2020, 47, 4659–4670. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y. GmWRKY17-mediated transcriptional regulation of GmDREB1D and GmABA2 controls drought tolerance in soybean. Plant Mol. Biol. 2023, 113, 157–170. [Google Scholar] [CrossRef]
- Banavath, J.N.; Chakradhar, T.; Pandit, V.; Konduru, S.; Guduru, K.K.; Akila, C.S.; Podha, S.; Puli, C.O.R. Stress Inducible Overexpression of AtHDG11 Leads to Improved Drought and Salt Stress Tolerance in Peanut (Arachis hypogaea L.). Front. Chem. 2018, 6, 34. [Google Scholar] [CrossRef]
- Zhao, J.P.; Su, X.H. Patterns of molecular evolution and predicted function in thaumatin-like proteins of Populus trichocarpa. Planta 2010, 232, 949–962. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Jami, S.K.; Swathi Anuradha, T.; Guruprasad, L.; Kirti, P.B. Molecular, biochemical and structural characterization of osmotin-like protein from black nightshade (Solanum nigrum). J. Plant Physiol. 2007, 164, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Deihimi, T.; Niazi, A.; Ebrahimi, M.; Kajbaf, K.; Fanaee, S.; Bakhtiarizadeh, M.R.; Ebrahimie, E. Finding the undiscovered roles of genes: An approach using mutual ranking of coexpressed genes and promoter architecture-case study: Dual roles of thaumatin like proteins in biotic and abiotic stresses. Springerplus 2012, 1, 30. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, M.; Zareie, R. Prediction, isolation, overexpression and antifungal activity analysis of Medicago truncatula var. truncatula putative thaumatin like proteins (TLP-1, -2, -3, -4 and -5). Turk. J. Biol. 2020, 44, 176–187. [Google Scholar] [CrossRef]
- Hayashi, M.; Shiro, S.; Kanamori, H.; Mori-Hosokawa, S.; Sasaki-Yamagata, H.; Sayama, T.; Nishioka, M.; Takahashi, M.; Ishimoto, M.; Katayose, Y.; et al. A thaumatin-like protein, Rj4, controls nodule symbiotic specificity in soybean. Plant Cell Physiol. 2014, 55, 1679–1689. [Google Scholar] [CrossRef]
- Singh, N.K.; Kumar, K.R.; Kumar, D.; Shukla, P.; Kirti, P.B. Characterization of a pathogen induced thaumatin-like protein gene AdTLP from Arachis diogoi, a wild peanut. PLoS ONE 2013, 8, e83963. [Google Scholar] [CrossRef] [PubMed]
- Vitali, A.; Pacini, L.; Bordi, E.; De Mori, P.; Pucillo, L.; Maras, B.; Botta, B.; Brancaccio, A.; Giardina, B. Purification and characterization of an antifungal thaumatin-like protein from Cassia didymobotrya cell culture. Plant Physiol. Biochem. 2006, 44, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakrabarti, C. Crystal structure analysis of NP24-I: A thaumatin-like protein. Planta 2008, 228, 883–890. [Google Scholar] [CrossRef]
- Min, K.; Ha, S.C.; Hasegawa, P.M.; Bressan, R.A.; Yun, D.J.; Kim, K.K. Crystal structure of osmotin, a plant antifungal protein. Proteins 2004, 54, 170–173. [Google Scholar] [CrossRef]
- Melchers, L.S.; Sela-Buurlage, M.B.; Vloemans, S.A.; Woloshuk, C.P.; Van Roekel, J.S.; Pen, J.; van den Elzen, P.J.; Cornelissen, B.J. Extracellular targeting of the vacuolar tobacco proteins AP24, chitinase and beta-1,3-glucanase in transgenic plants. Plant Mol. Biol. 1993, 21, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Y.; Wang, H.X.; Ng, T.B. First chromatographic isolation of an antifungal thaumatin-like protein from French bean legumes and demonstration of its antifungal activity. Biochem. Biophys. Res. Commun. 1999, 263, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Samac, D.A.; Penuela, S.; Schnurr, J.A.; Hunt, E.N.; Foster-Hartnett, D.; Vandenbosch, K.A.; Gantt, J.S. Expression of coordinately regulated defence response genes and analysis of their role in disease resistance in Medicago truncatula. Mol. Plant Pathol. 2011, 12, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Otte, O.; Pachten, A.; Hein, F.; Barz, W. Early elicitor-induced events in chickpea cells: Functional links between oxidative burst, sequential occurrence of extracellular alkalinisation and acidification, K+/H+ exchange and defence-related gene activation. Z. Naturforsch. C J. Biosci. 2001, 56, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Benko-Iseppon, A.M.; Winter, P.; Huettel, B.; Staginnus, C.; Muehlbauer, F.J.; Kahl, G. Molecular markers closely linked to fusarium resistance genes in chickpea show significant alignments to pathogenesis-related genes located on Arabidopsis chromosomes 1 and 5. Theor. Appl. Genet. 2003, 107, 379–386. [Google Scholar] [CrossRef]
- Trudel, J.; Grenier, J.; Potvin, C.; Asselin, A. Several thaumatin-like proteins bind to beta-1,3-glucans. Plant Physiol. 1998, 118, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Grenier, J.; Potvin, C.; Trudel, J.; Asselin, A. Some thaumatin-like proteins hydrolyse polymeric beta-1,3-glucans. Plant J. 1999, 19, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Petre, B.; Major, I.; Rouhier, N.; Duplessis, S. Genome-wide analysis of eukaryote thaumatin-like proteins (TLPs) with an emphasis on poplar. BMC Plant Biol. 2011, 11, 33. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Petre, B. Toward the Discovery of Host-Defense Peptides in Plants. Front. Immunol. 2020, 11, 1825. [Google Scholar] [CrossRef]
- Jayaprakash, A.; Roy, A.; Thanmalagan, R.R.; Arunachalam, A.; Ptv, L. Immune response gene coexpression network analysis of Arachis hypogaea infected with Aspergillus flavus. Genomics 2021, 113, 2977–2988. [Google Scholar] [CrossRef]
- Kang, J.N.; Lee, W.H.; Won, S.Y.; Chang, S.; Hong, J.P.; Oh, T.J.; Lee, S.M.; Kang, S.H. Systemic Expression of Genes Involved in the Plant Defense Response Induced by Wounding in Senna tora. Int. J. Mol. Sci. 2021, 22, 10073. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sui, X.; Wang, Y.; Zhu, M.; Zhou, Y.; Gao, F. Genome-Wide Analyses of Thaumatin-like Protein Family Genes Reveal the Involvement in the Response to Low-Temperature Stress in Ammopiptanthus nanus. Int. J. Mol. Sci. 2023, 24, 2209. [Google Scholar] [CrossRef] [PubMed]
- Fecht-Christoffers, M.M.; Braun, H.P.; Lemaitre-Guillier, C.; VanDorsselaer, A.; Horst, W.J. Effect of manganese toxicity on the proteome of the leaf apoplast in cowpea. Plant Physiol. 2003, 133, 1935–1946. [Google Scholar] [CrossRef]
- Del Campillo, E.; Lewis, L.N. Identification and kinetics of accumulation of proteins induced by ethylene in bean abscission zones. Plant Physiol. 1992, 98, 955–961. [Google Scholar] [CrossRef]
- Piromyou, P.; Nguyen, H.P.; Songwattana, P.; Boonchuen, P.; Teamtisong, K.; Tittabutr, P.; Boonkerd, N.; Alisha Tantasawat, P.; Gottfert, M.; Okazaki, S.; et al. The Bradyrhizobium diazoefficiens type III effector NopE modulates the regulation of plant hormones towards nodulation in Vigna radiata. Sci. Rep. 2021, 11, 16604. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, Z.; Yin, W.; Xu, K.; Wang, S.; Shang, Q.; Sa, W.; Liang, J.; Wang, L. Genome-wide analysis of the Thaumatin-like gene family in Qingke (Hordeum vulgare L. var. nudum) uncovers candidates involved in plant defense against biotic and abiotic stresses. Front. Plant Sci. 2022, 13, 912296. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fukuda, M.; Igarashi, D.; Sunaoshi, M. Cytokinin-binding proteins from tobacco callus share homology with osmotin-like protein and an endochitinase. Plant Cell Physiol. 2000, 41, 148–157. [Google Scholar] [CrossRef][Green Version]
- Ding, L.N.; Li, Y.T.; Wu, Y.Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.L. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef] [PubMed]
- Yamchi, A.; Ben, C.; Rossignol, M.; Zareie, S.R.; Mirlohi, A.; Sayed-Tabatabaei, B.E.; Pichereaux, C.; Sarrafi, A.; Rickauer, M.; Gentzbittel, L. Proteomics analysis of Medicago truncatula response to infection by the phytopathogenic bacterium Ralstonia solanacearum points to jasmonate and salicylate defence pathways. Cell. Microbiol. 2018, 20, e12796. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lan, L.; Jin, Y.; Yu, N.; Wang, D.; Wang, E. Mechanisms underlying legume-rhizobium symbioses. J. Integr. Plant Biol. 2022, 64, 244–267. [Google Scholar] [CrossRef] [PubMed]
- Berrabah, F.; Ratet, P.; Gourion, B. Legume Nodules: Massive Infection in the Absence of Defense Induction. Mol. Plant Microbe Interact. 2019, 32, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Hochholdinger, F.; Li, C. Comparative analyses of three legume species reveals conserved and unique root extracellular proteins. Proteomics 2012, 12, 3219–3228. [Google Scholar] [CrossRef]
- Regalado, A.P.; Ricardo, C.P. Study of the intercellular fluid of healthy Lupinus albus organs. Presence of a chitinase and a thaumatin-like protein. Plant Physiol. 1996, 110, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Hanselle, T.; Ichinoseb, Y.; Barz, W. Biochemical and molecular biological studies on infection (Ascochyta rabiei)-induced thaumatin-like proteins from chickpea plants (Cicer arietinum L.). Z. Naturforsch. C J. Biosci. 2001, 56, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Orona-Tamayo, D.; Wielsch, N.; Escalante-Perez, M.; Svatos, A.; Molina-Torres, J.; Muck, A.; Ramirez-Chavez, E.; Adame-Alvarez, R.M.; Heil, M. Short-term proteomic dynamics reveal metabolic factory for active extrafloral nectar secretion by Acacia cornigera ant-plants. Plant J. 2013, 73, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Toyotake, Y.; Cho, H.N.; Kawamoto, J.; Kurihara, T. A novel 1-acyl-sn-glycerol-3-phosphate O-acyltransferase homolog for the synthesis of membrane phospholipids with a branched-chain fatty acyl group in Shewanella livingstonensis Ac10. Biochem. Biophys. Res. Commun. 2018, 500, 704–709. [Google Scholar] [CrossRef]
- Alonso, A.; Pulido, R. The extended human PTPome: A growing tyrosine phosphatase family. FEBS J. 2016, 283, 2197–2201. [Google Scholar] [CrossRef]
- Chuong, N.N.; Hoang, X.L.T.; Nghia, D.H.T.; Dai, T.N.T.; Thi, V.L.; Thao, N.P. Protein Phosphatase Type 2C Functions in Phytohormone-Dependent Pathways and in Plant Responses to Abiotic Stresses. Curr. Protein Pept. Sci. 2021, 22, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Berndsen, C.E.; Wolberger, C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014, 21, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lee, M.J.; Solis, N.V.; Phan, Q.T.; Swidergall, M.; Ralph, B.; Ibrahim, A.S.; Sheppard, D.C.; Filler, S.G. Aspergillus fumigatus CalA binds to integrin α5β1 and mediates host cell invasion. Nat. Microbiol. 2016, 2, 16211. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Detchemendy, T.W.; Pajerowska-Mukhtar, K.M.; Mukhtar, M.S. NPR1 in JazzSet with Pathogen Effectors. Trends Plant Sci. 2018, 23, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, Y.; Cai, H.; Guo, M.; Chai, M.; She, Z.; Ye, L.; Cheng, Y.; Wang, B.; Qin, Y. The bZIP Transcription Factor GmbZIP15 Negatively Regulates Salt- and Drought-Stress Responses in Soybean. Int. J. Mol. Sci. 2020, 21, 7778. [Google Scholar] [CrossRef] [PubMed]
- Sornaraj, P.; Luang, S.; Lopato, S.; Hrmova, M. Basic leucine zipper (bZIP) transcription factors involved in abiotic stresses: A molecular model of a wheat bZIP factor and implications of its structure in function. Biochim. Biophys. Acta 2016, 1860, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, Y.; Li, Z.; She, Z.; Chai, M.; Aslam, M.; He, Q.; Huang, Y.; Chen, F.; Chen, H.; et al. The bZIP transcription factor GmbZIP15 facilitates resistance against Sclerotinia sclerotiorum and Phytophthora sojae infection in soybean. Iscience 2021, 24, 102642. [Google Scholar] [CrossRef] [PubMed]
- Fierens, E.; Gebruers, K.; Voet, A.R.; De Maeyer, M.; Courtin, C.M.; Delcour, J.A. Biochemical and structural characterization of TLXI, the Triticum aestivum L. thaumatin-like xylanase inhibitor. J. Enzym. Inhib. Med. Chem. 2009, 24, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Smole, U.; Bublin, M.; Radauer, C.; Ebner, C.; Breiteneder, H. Mal d 2, the thaumatin-like allergen from apple, is highly resistant to gastrointestinal digestion and thermal processing. Int. Arch. Allergy Immunol. 2008, 147, 289–298. [Google Scholar] [CrossRef]
- He, L.; Li, L.; Zhu, Y.; Pan, Y.; Zhang, X.; Han, X.; Li, M.; Chen, C.; Li, H.; Wang, C. BolTLP1, a Thaumatin-like Protein Gene, Confers Tolerance to Salt and Drought Stresses in Broccoli (Brassica oleracea L. var. Italica). Int. J. Mol. Sci. 2021, 22, 11132. [Google Scholar] [CrossRef]
- Iquebal, M.A.; Soren, K.R.; Gangwar, P.; Shanmugavadivel, P.S.; Aravind, K.; Singla, D.; Jaiswal, S.; Jasrotia, R.S.; Chaturvedi, S.K.; Singh, N.P.; et al. Discovery of Putative Herbicide Resistance Genes and Its Regulatory Network in Chickpea Using Transcriptome Sequencing. Front. Plant Sci. 2017, 8, 958. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Au, T.K.; Lam, T.L.; Ye, X.Y.; Wan, D.C. Inhibitory effects of antifungal proteins on human immunodeficiency virus type 1 reverse transcriptase, protease and integrase. Life Sci. 2002, 70, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Zemanek, E.C.; Wasserman, B.P. Issues and advances in the use of transgenic organisms for the production of thaumatin, the intensely sweet protein from Thaumatococcus danielli. Crit. Rev. Food Sci. Nutr. 1995, 35, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Faus, I. Recent developments in the characterization and biotechnological production of sweet-tasting proteins. Appl. Microbiol. Biotechnol. 2000, 53, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewski, G.; Niedziela, A.; Szwacka, M.; Szczytt, K.N. Modification of tomato taste in transgenic plants carrying a thaumatin gene from Thaumatococcus Daniellii Benth. Plant Breed. 2003, 122, 347–351. [Google Scholar] [CrossRef]
- Masuda, T.; Kitabatake, N. Developments in biotechnological production of sweet proteins. J. Biosci. Bioeng. 2006, 102, 375–389. [Google Scholar] [CrossRef]




| Interactions | Predicted Interaction Partners | Protein Accessions |
|---|---|---|
| Direct interaction with AhTLP1 | TYR_PHOSPHATASE_2 domain-containing protein | Ahy_A09g042676 |
| Protein phosphatase 2C and cyclic nucleotide-binding/kinase domain-containing protein | Ahy_A05g023223, Ahy_B05g074771 | |
| PlsC domain-containing protein | Ahy_B10g101660, Ahy_A07g037205 | |
| 1-acyl-sn-glycerol-3-phosphate acyltransferase | Ahy_A07g031308 | |
| 3-deoxy-d-manno-octulosonic-acid transferase | Ahy_A02g009900 | |
| E3 ubiquitin–protein ligase | Ahy_B06g084168 | |
| Uncharacterized protein | Ahy_A02g009985, Ahy_B09g095505 | |
| Indirect interaction with AhTLP1 | Rac-like GTP-binding protein RAC13 | Ahy_B10g101133 |
| Ribonucleoside–diphosphate reductase subunit beta | Ahy_B08g089004 | |
| Clathrin heavy chain | Ahy_B07g088015, Ahy_B09g094835, Ahy_B07g086972, Ahy_B10g103140 | |
| Beta-adaptin-like protein | Ahy_B10g105006 | |
| FYVE-type domain-containing protein | Ahy_B08g090320 | |
| Subunit of the adaptor protein complex 2 | Ahy_B08g093980 | |
| Beta-adaptin-like protein | Ahy_B08g091239 |
| Interactions | Predicted Interaction Partners | Protein Accessions |
|---|---|---|
| Direct interaction with GmOLPa | Sucrose–phosphatase 2 | A0A0R0I3G3, A0A0R0E8K1 |
| Bet_v_1 domain-containing protein | I1LP68_SOYBN | |
| Regulatory protein NPR1 | A0A0R0IBM0 | |
| Sucrose–phosphatase 1 | I1L9P5_SOYBN | |
| Ribonucleoprotein | C6TEA3_SOYBN | |
| UBIQUITIN_CONJUGAT_2 domain-containing protein | I1K9A6_SOYBN | |
| Calmodulin-like protein 1 | C6SWC0_SOYBN, C6TAX4_SOYBN | |
| ABC transporter family protein | K7LE00_SOYBN | |
| Indirect interaction with GmOLPa | Glycosyl hydrolase 31 family | I1MG74_SOYBN, A0A0R0I6Y6 |
| Sucrose–phosphate synthase 2 | I1KY06_SOYBN, I1M6Y2_SOYBN, I1N0X9_SOYBN, I1KFX9_SOYBN, K7M052_SOYBN, I1MU40_SOYBN | |
| Peroxisome biogenesis protein | I1M423_SOYBN, I1LU78_SOYBN |
| Interactions | Predicted Interaction Partners | Protein Accessions |
|---|---|---|
| Direct interaction with GmOLPb | Cytochrome P450 family protein | K7MVY7_SOYBN, A0A0R0GJG3, I1N5Y9_SOYBN, K7MVY4_SOYBN, A0A0R0EGW1, I1JDB7_SOYBN |
| Trypsin inhibitor A | I1N2Q3_SOYBN | |
| Fe2+/Zn2+ regulated transporter | IRT | |
| Sucrose–phosphatase 1 | I1L9P5_SOYBN | |
| bZIP transcription factor | GmbZIP1 | |
| Indirect interaction with GmOLPb | Tim44 domain-containing protein | I1JVJ8_SOYBN, K7LB35_SOYBN |
| Sucrose–phosphate synthase 2 | I1KY06_SOYBN, I1KFX9_SOYBN, I1M6Y2_SOYBN, K7M052_SOYBN, I1N0X9_SOYBN, I1MU40_SOYBN | |
| Glycosyl hydrolase 31 family | A0A0R0I6Y6, I1MG74_SOYBN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, L.; Wei, S.; Li, Y. Thaumatin-like Proteins in Legumes: Functions and Potential Applications—A Review. Plants 2024, 13, 1124. https://doi.org/10.3390/plants13081124
Feng L, Wei S, Li Y. Thaumatin-like Proteins in Legumes: Functions and Potential Applications—A Review. Plants. 2024; 13(8):1124. https://doi.org/10.3390/plants13081124
Chicago/Turabian StyleFeng, Lanlan, Shaowei Wei, and Yin Li. 2024. "Thaumatin-like Proteins in Legumes: Functions and Potential Applications—A Review" Plants 13, no. 8: 1124. https://doi.org/10.3390/plants13081124
APA StyleFeng, L., Wei, S., & Li, Y. (2024). Thaumatin-like Proteins in Legumes: Functions and Potential Applications—A Review. Plants, 13(8), 1124. https://doi.org/10.3390/plants13081124